Детальные исследования структуры хромосом в различные периоды их цикла в митозе и мейозе, а также в некоторых неделящихся клетках позволили выявить важные особенности тонкой микроскопической и субмикроскопической организации хромосом.
Хромонемы
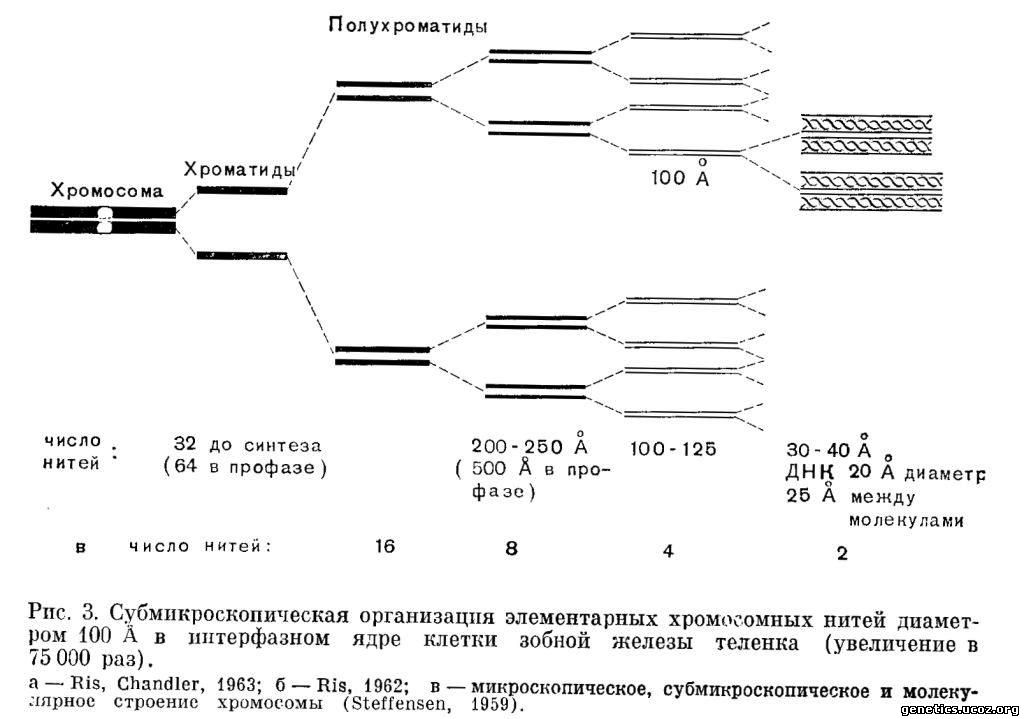
Хромосомы активно участвуют в процессах внутриклеточного обмена и в связи с биохимическими и физиологическими изменениями, происходящими в клетке в процессах ее деления, дифференцировки и функционирования, они претерпевают сложный цикл морфологических и физико-химических преобразований. Основу микроскопического строения хромосом во всех стадиях этого цикла составляет одна или несколько спирализованных дезоксирибоиуклеопротеидпых нитей — хромонем (рис. 2, в). Одной из проблем цитогенетики является выяснение вопроса, сколько таких нитевидных структур образуют хромосому. Эта проблема менее важна для генетика, так как для него функциональной единицей хромосомы в клеточном делении, в мутациях, в разъединении генов и в кроссинговере всегда является хроматида. Многочисленными цитогенетическими исследованиями показано, что хроматида в свою очередь состоит из субъединиц.
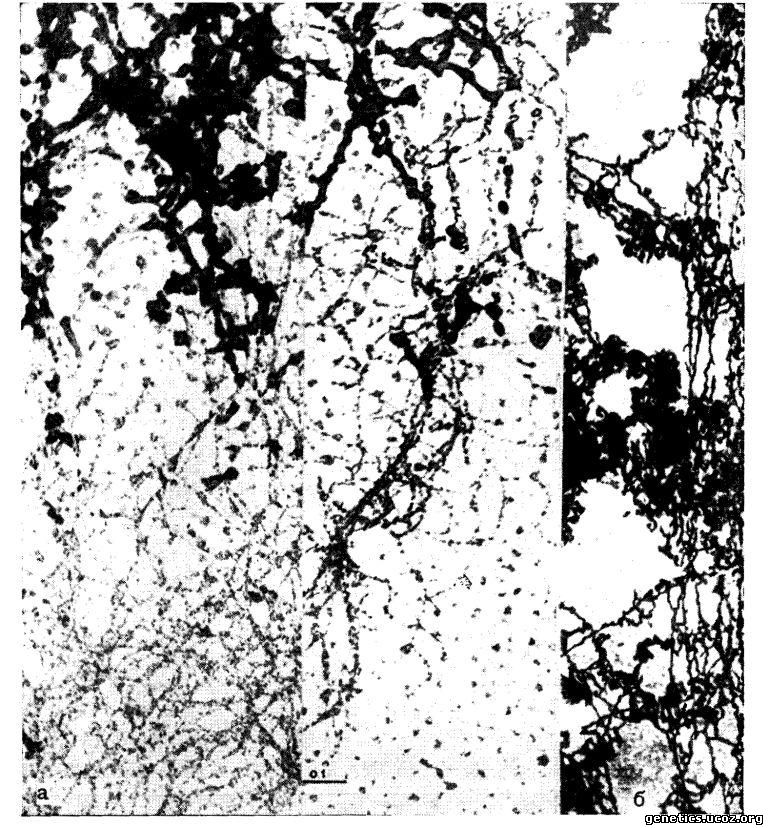
Электронномикроскопические исследования устанавливают, что хромосома представляет собой пучок из 8 —16 элементарных ДНК-протеидных нитей, каждая из которых имеет диаметр 100—150 А (рис. 3, а, б). Элементарные хромосомные нити образованы одной (двойной) молекулой ДНК, соединенной с гистонным белком (Ris, Chandler, 1963) (рис. 3, в).
Хромомеры
Данные генетического анализа устанавливают, что хромосома дифференцирована по длине на специфические участки, влияющие па характер развития клеток и организма, — гены.
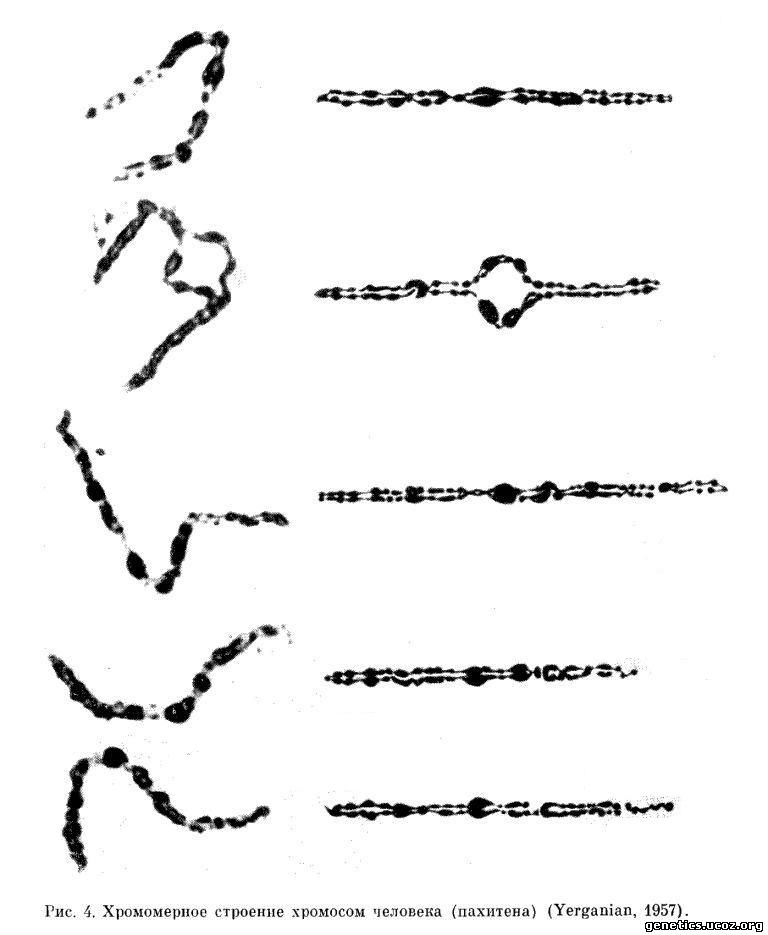
Функциональной дифференциации хромосомы по длине соответствует структурная дифференциация, выражающаяся на микроскопическом уровне в хромомерном строении. Последнее наиболее отчетливо наблюдается в ранней профазе мейоза, когда хромосомы имеют вид тончайших нитей с утолщениями — хромомерами. Различаясь между собой по величине, форме, содержанию ДНК и положению в хромосоме, хромомеры обладают отчетливо выраженной индивидуальностью и придают хромосомам и отдельным их участкам постоянный и определенный рисунок, строго фиксированный наследственно. Это позволило для ряда организмов, включая человека, составить карты пахитенных хромосом (рис. 4; рис. 64—65).
Центромера
Хромосома может функционировать в митозе и мейозе только тогда, когда она обладает центромерой (кинетическое тельце, кинетохор). Вместе с центриолью в цитоплазме кинетическое тельце хромосомы организует белковые мицеллы цитоплазмы, ориентируя их для построения ахроматинового веретена.
Именно этот район хромосомы включается в действие специфических сил отталкивания хромосом от обоих полюсов клетки, определяя расположение хромосом по экватору веретена в метафазе. Отталкивание центромерных районов одного от другого определяет расположение всех хромосом в метафазе на равных расстояниях между собой, а отталкивание центромерных районов сестринских хроматид является одним из факторов, определяющих их расхождение к противоположным полюсам в анафазе (Е. Н. Герасимова-Навашина, 1951). Таким образом, вся динамика хромосом в митозе и мейозе определяется функционированием центромеры. Участок хромосомы, лишенный центромеры, — ацентрический фрагмент — при первом же митозе не может принимать в нем участия, остается пассивным, не включается в сестринские ядра и резорбируется в цитоплазме.
Таким образом, судьба хромосом после действия на них ионизирующей радиации или химических мутагенов полностью определяется поведением их центроморных районов. Поэтому исследование строения и свойств центромеры приобретает в настоящее время особое значение.
Обычно центромерный участок представлен в теле компактной, интенсивно окрашивающейся метафазной хромосомы неокрашивающимся перерывом: первичной перетяжкой между двумя плечами хромосомы (см. рис. 2). Детальными исследованиями установлено, что этот участок содержит хромопемы с 2—3 парами хромомер (Lima-de-Faria, 1949, 1956).
В результате действия рентгеновых лучей в районе центромеры может произойти разрыв под прямым углом к продольной оси хромосомы, в результате чего возникают две хромосомы с фрагментированными центромерами. Каждая такая центромера соединена с двумя хроматидами. Эта структура, развертываясь, дает начало изохромосомам (Darlington, 1939; Lima-de-Faria, 1956), имеющим два полностью идентичных плеча. Таким образом, было установлено сложное строение центромеры, отдельные элементы которой могут функционировать в митозе после ее разрыва (Swanson, 1960) (см. рис. 20).
Теломеры
На каждом из свободных концов хромосома содержит специальную структуру— теломеру. По данным Lima-de-Faria и Sarvella (1958), теломера хромосом ряда растений (Scilla, Solanum, Secale и др.) имеет сложное строение: в ее состав входит до 8 хромомер, которые могут быть обнаружены микроскопически. Теломера обладает отчетливо выраженными свойствами, обнаруженными при исследовании хромосом, поврежденных ионизирующей радиацией: свободный конец хромосомы с теломерой не способен соединяться с другими хромосомами или фрагментами, в то время как конец хромосомы, лишенный (в результате разрывов) теломеры, оказывается «ненасыщенным», делает хромосому нестойкой и присоединяется к таким же разорванным концам других хромосом. Если же хромосома в момент разрыва образована двумя хроматидами, их оторванные концы соединяются между собой, образуя днцентрическую хроматиду. Разорванные концы хромосом могут иногда «залечиваться», но подобные случаи редки (McClintock, 1941).
Матрикс
Как предполагают многие цитологи, хромонемы в хромосоме окружены ахроматическим веществом — матриксом и поверхностной оболочкой (см. рис. 2, в). Матрикс был обнаружен рядом цитологов на хромосомах кукурузы, дрозофилы и его реальность постулируется в современных крупных монографиях по цитологии (Swanson, 1960). Полагают, что матрикс содержит ДНК. Матрикс образован, по-видимому, продуктами активности хромосомы, концентрирующимися на ее поверхности при спирализации хромосомы. Об этом свидетельствуют данные радиоавтографического анализа хромосом в стадии диплотены у тритона (Gall, Callan, 1962).
Вторичные перетяжки и ядрышко
Открытие закономерной связи между ядрышками и определенными хромосомами принадлежит С. Г. Навашину (1912), установившему, что в профазе митоза у гальтонии ядрышки всегда связаны со спутничными хромосомами.
Heitz (1931) показал, что ядрышки возникают па нити, соединяющей спутник с телом хромосомы. Дальнейшими исследованиями эти наблюдения подтверждены.
Вторичные перетяжки на метафазных хромосомах обычно представляют собой неспирализованный участок хромюнем, вокруг которого ранее было сформировано ядрышко. После исчезновения ядрышка (в профазе) в участке его локализации остается тонкая нить или слабо красящийся перерыв в хромосоме. Спутники представляют собой участки хромосомы, расположенные дистальнее вторичной перетяжки. В телофазе в период реорганизации ядра ядрышко вновь формируется на идентичных участках сестринских хромосом.
У многих растительных организмов ядрышко формируется нитью или прилежащим к нити участком спутничной хромосомы; число ядрышек соответствует числу спутничных хромосом и отношение величин ядрышек, содержащихся в ядре, соответствует отношению длин нитей (вторичных перетяжек) спутничных хромосом (Heitz, 1931).
Ядрышки могут образовываться также в районе вторичной перетяжки, не обязательно представляющей собой спутничную нить.
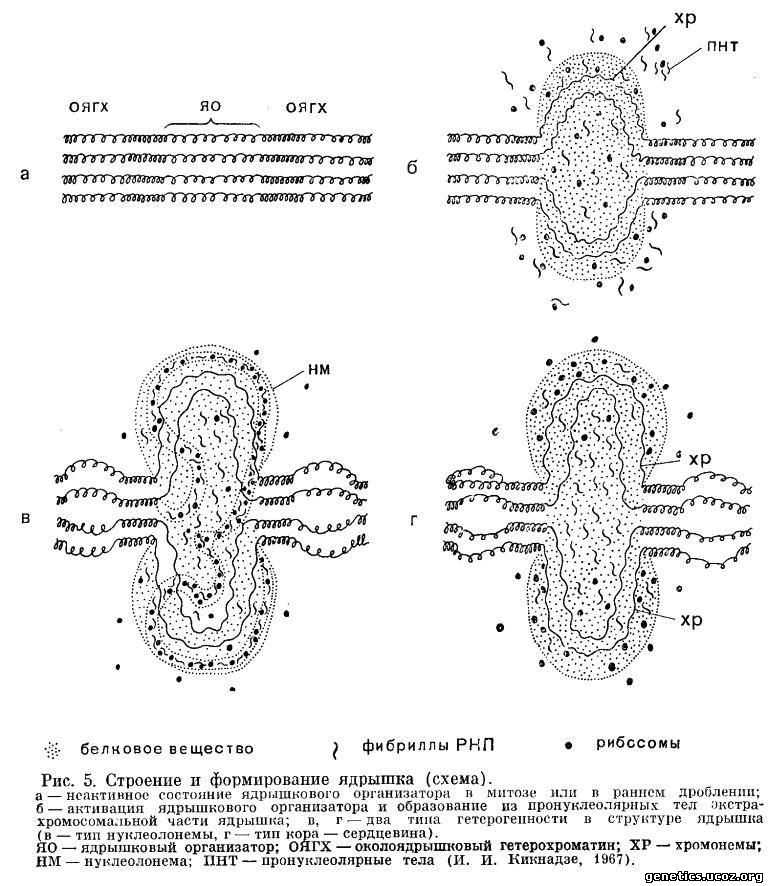
Природа и функции ядрышка длительное время оставались загадочными, и только в последнее десятилетие был пролит свет на эти вопросы (см. обзоры И. И. Кикнадзе и Е. Беляевой, 1967). В настоящее время точно установлено, что ядрышко представляет собой видоизмененный специализированный участок хромосомы, сформированный деспирализованными хромонемами определенного локуса хромосомы — ядрышкового организатора (рис. 5).
Кроме деспирализованных хромонем, ядрышко содержит сильно развитую и сложно организованную экстрахромосомную часть, образованную продуктами деятельности ядрышкового организатора. Эта часть ядрышка представляет собой гетерогенное образование, состоящее из фибрилл, гранул и аморфного вещества. Генетическая роль ядрышка выяснена исследованиями молекулярной биологии. Установлено, что функция ядрышка состоит в синтезе рибосомальной РНК и в нем осуществляются определенные этапы формирования рибосом (Г. П. Георгиев, 1964; И. И. Кикнадзе и Е. Беляева, 1967).
В ядрышке синтезируется 80% РНК клетки. В исследованиях с использованием метода гибридизации молекул РНК — ДНК показано, что ядрышковый организатор хромосомы имеет полигенную природу (Г. П. Георгиев,1964; Ritossa, Spiegelman, 1965). Обычно в нормальных условиях функционирует определенное для клеток данного вида организма и данной стадии развития количество ядрышковых организаторов. Усиление функциональной активности клетки сопровождается активизацией деятельности этих организаторов или же вступлением в действие дополнительных, до того неактивных организаторов.