Наиболее обстоятельные исследования по анализу характера повреждений хромосом, вызываемых действием вируса, проведены с вирусом кори. При анализе хромосом в культуре лейкоцитов периферической крови больных и субъектов, вакцинированных живым аттенуированным вирусом кори (Nichols и др., 1962, 1963), было установлено, что основной тип повреждений состоит в одиночных разрывах, относящихся к категории «замедленных изолокусных разрывов». Этот тип разрывов был ранее установлен и обозначен этим термином на хромосомах растений (Ostergren, Wakonic, 1954) и обусловлен возникновением (или проявлением) в хромосоме лабильного локуса. Предполагается, что лабильный локус возникает в хромосоме, тогда когда она представляет собой функционально одну нить. В период удвоения хромосомы этот локус также редуплицируется, и таким образом каждая из двух хроматид может содержать лабильный локус. В следующем митозе в хромосоме обнаруживается серия разнообразных аномалий, включая появление просветов и перетяжки в одной хроматиде и разрыв в соответствующем участке второй хроматиды; полный разрыв в обоих хроматидах наблюдается значительно реже, перетяжки — в обоих или лишь в одной хроматиде. Эти вариации в проявлении повреждения, как полагают авторы, могут быть обусловлены тем моментом в митозе, когда лабильный локус разорвался (Nichols и др., 1964). Характерной особенностью разрывов хромосом, вызванных действием вируса кори, является, как правило, отсутствие воссоединений и хромосомных перестроек. Этим вирусные повреждения хромосом отчетливо отличаются от повреждений, возникающих в результате действия ионизирующей радиации и некоторых химических мутагенов.
При анализе повреждений хромосом под действием вируса кори на четыре различные культуры клеток человека был обнаружен феномен, не наблюдавшийся при исследовании культуры лейкоцитов больных корью (Nichols и др., 1965).
В этих культурах, кроме одиночных разрывов хромосом, типичных для хромосомных наборов у больных корью, были обнаружены многочисленные случаи крайней степени фрагментации всех хромосом до распыления их на мельчайшие хромосомные участки (рис. 96). Этот весьма специфический для действия вируса кори феномен повреждения хромосом авторы обозначили специальным термином «распыление» (pulverization).
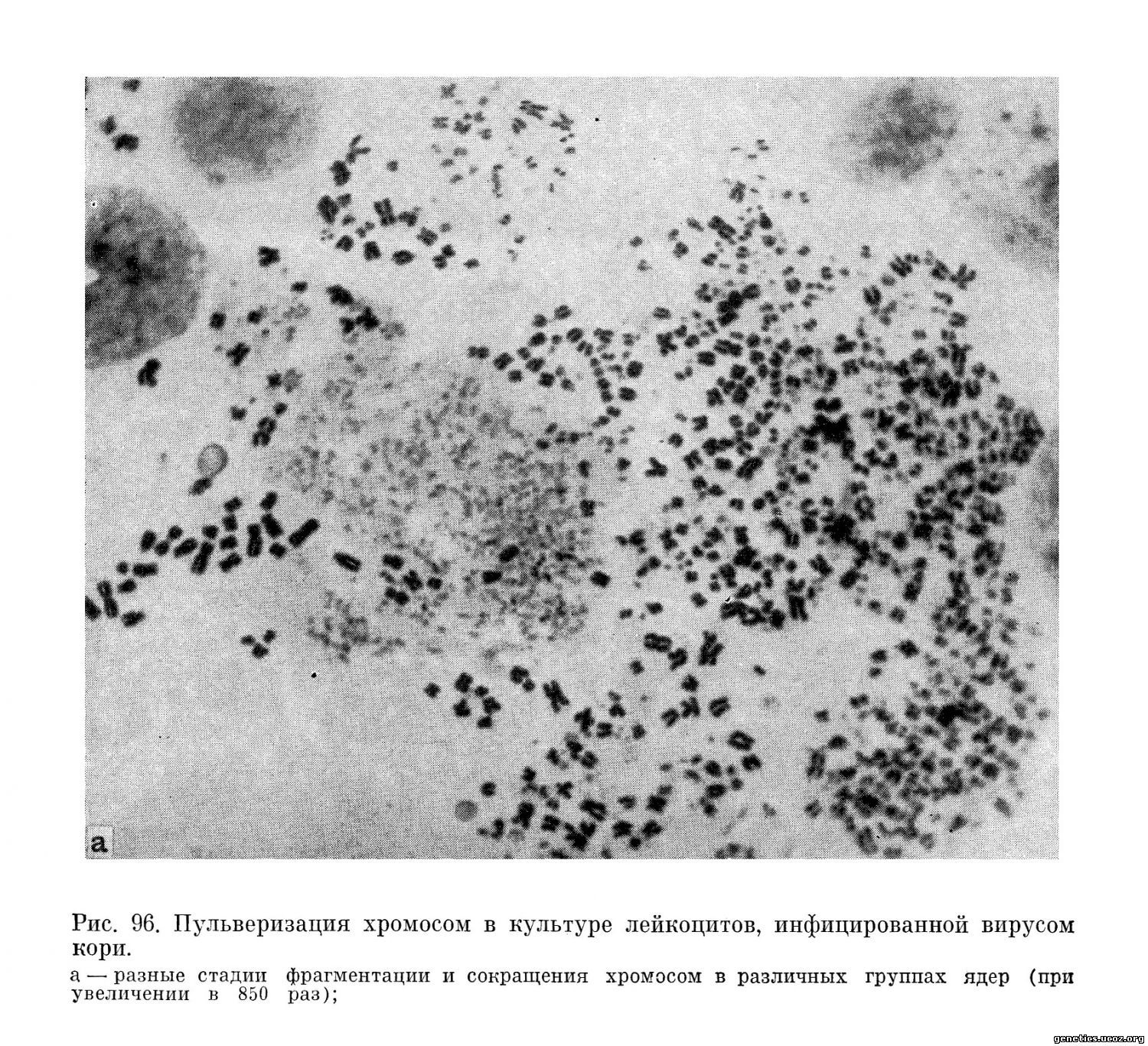
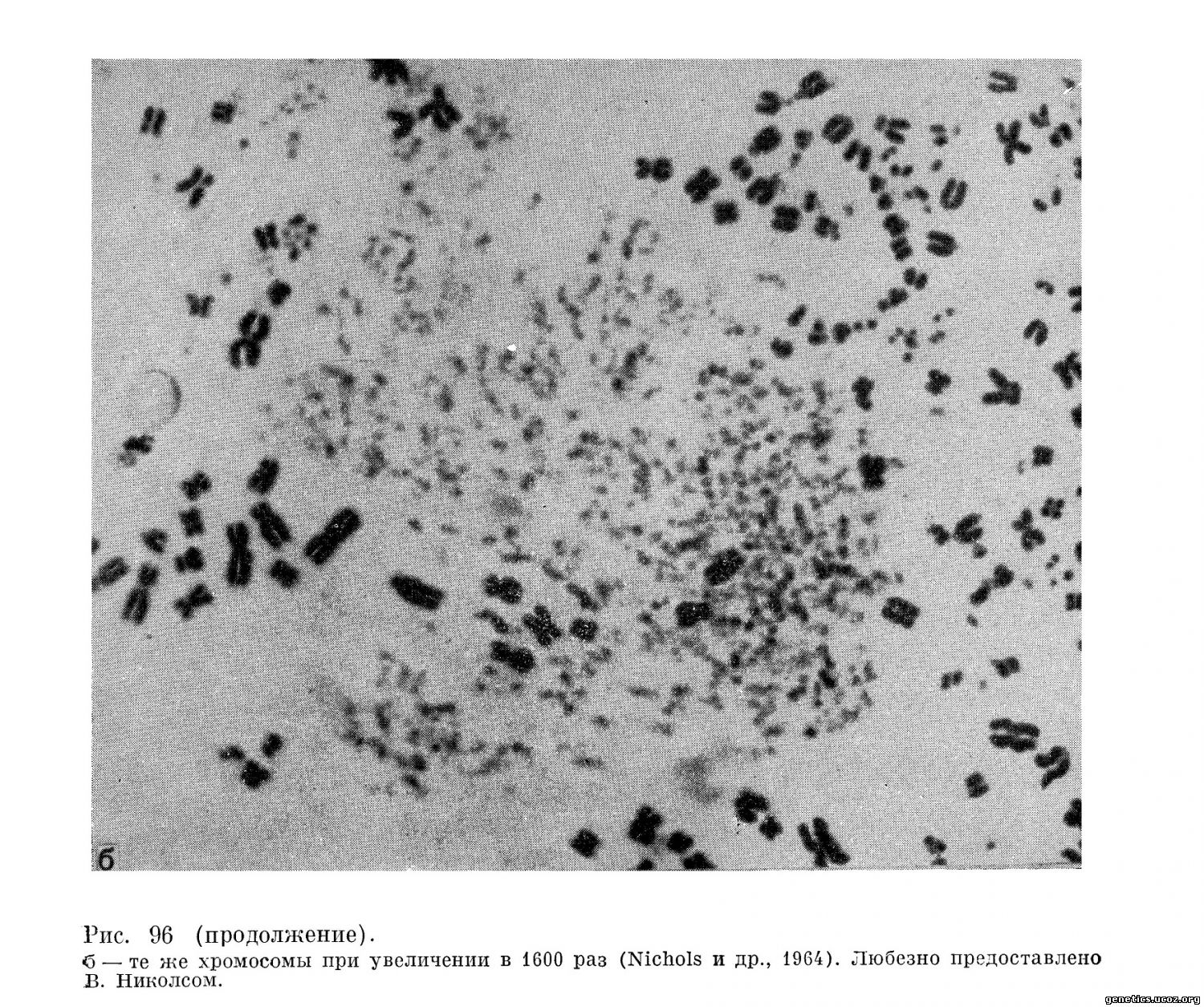
На этих же культурах как результат действия вируса кори наблюдалась тенденция клеток сливаться и формировать общие синцитии с многочисленными хромосомными наборами; для них особенно характерно «распыление» хромосом. Обращает на себя внимание тот факт, что каждая группа хромосом имеет собственный, характерный рисунок распыления.
Анализ частоты клеток с различным числом единичных разрывов хромосом (0—7) и клеток с «распылением» хромосом показал, что клетки с единичными разрывами распределяются случайно, согласно пуассоновскому распределению, а клетки с «распыленными» хромосомами принадлежат к другой статистической популяции.
Таким образом, «распыление» хромосом представляет собой особый тип вирусного поражения их, а не результат аккумуляции в ядре множественных единичных разрывов.
К феномену распыления хромосом можно отнести случаи крайне сильной фрагментации хромосом в культуре эмбрионального легкого человека, инфицированной вирусом герпеса (Stich и др., 1964). Сходное поражение хромосом, выражающееся в чрезвычайно высокой степени их фрагментации, обнаружено у пациентов, вакцинированных против желтой лихорадки живым аттенуированным вирусом (Hamden, 1964) в культурах клеток человека, инфицированных вирусом истинной чумы птиц и болезни Ньюкасла (В. В. Башкова, 1966), вирусом Сендай (Aula, Saksela, 1966) и аденовирусом 12 (Stich и др., 1964).
На основании детального исследования действия вируса кори на хромосомы человека in vivo и in vitro Aula (1965) пришел к заключению, что морфологически повреждения хромосом в обоих случаях идентичны и представляют собой в основном хроматидные разрывы и хроматидные просветы (gaps) без тенденции разорванных участков к воссоединению или перестройкам. Этот же тип повреждения хромосом установлен и при инфицировании культуры клеток человека и животных другими вирусами: герпес простой, SV-40, полиома, серозный гепатит, аденовирус 12, ветряная оспа, асептический менингит (Hampar, Ellison, 1963; Mazzone, Yerganian, 1963; Stich и др., 1964; Koprowski и др., 1962; Yerganian и др., 1962; Shein, Enders, 1963; Vogt, Dulbecco, 1963; Cooper, Black, Ponten и др., 1963; Wolman и др., 1964; Moorhead, Saksela, 1965; Aya, Makino, 1966; Makino и др., 1966).
Gripenberg (1965) детально исследовала особенности повреждения хромосом в 2 случаях вирусной инфекции: у пациента с exanthema subitum и пациента с ветряной оспой (рис. 97).

Оба этих случая сопровождались высокой частотой клеток с разрывами хромосом (54 и 57% соответственно). Хотя частота клеток с разрывами в обоих случаях была сходной, однако цитологическая картина повреждений хромосом отчетливо различалась. В случае действия вируса exanthema subitum в хромосомах пациента появлялись перетяжки и хроматидные разрывы. В случае ветряной оспы проявления поражений хромосом были значительно более разнообразны: перерывы в хромосомах (gaps) шире, более выражена тенденция к полным разрывам, очень часто появляются фрагменты изохроматидного типа, встречаются перестройки, обусловленные обменом участками хромосом. Трудно решить, вызваны ли эти различия разной степенью поражения хромосом или же они представляют собой различные стадии — более раннюю и более позднюю — одного и того же процесса.
В обстоятельной работе Moorhead и Saksela (1965) установили последовательность, в которой возникают разные типы повреждений хромосомных наборов в диплоидной культуре фибробластов человека (линия 1—38), инфицированной SV-40 (на 9-м и 30-м пассажах этой культуры).
Установлено, что морфологическая трансформация клеток этой культуры под действием SV-40 происходит после ряда пассажей: между 8-й и 11-й неделей после инфицирования (Ponten и др., 1963). В соответствии с этими данными находятся и наблюдения Moorhead и Saksela: единичные клетки с измененными хромосомами обнаружены через 3—7 недель после инфицирования культуры, и только через 16 недель после инфицирования (через 7—10 пассажей) частота хромосомных аберраций резко возрастает. При этом обнаруживается закономерная смена типов повреждений хромосомных наборов: вначале повышается частота некрасящихся просветов (gaps), затем хроматидных разрывов; одновременно повышается частота тетраплоидных клеток, затем появляются как следствие вторичные повреждения хромосом — хромосомные перестройки, сопровождающиеся различными типами аберраций, — дицентрики, кольца, фрагменты и, наконец, сильно повышается частота тетраплоидных и анэуплоидных клеток с числами хромосом от 76 до 85, содержащих многочисленные хромосомные аберрации. Moorhead и Saksela не дали в своих исследованиях ответа на вопрос, почему хроматидные разрывы, указывающие на первичное повреждение хромосом в процессе синтеза ДНК, появляются при инфицировании культуры ранних пассажей (9) так поздно — через 7—12 недель после инфицирования. Инфицирование более старой культуры (30-го пассажа) в их опытах сопровождалось более ранним появлением клеток с поврежденными хромосомами и более ранней морфологической трансформацией культуры.
Сходные данные были получены ранее при инфицировании вирусом SV-40 культуры фибробластов человека, находящейся в III фазе роста (старение). В этих условиях разнообразные изменения хромосомных наборов обнаруживались уже в течение первых 2—4 дней: вначале повышалась частота разрывов хроматид и хромосом, затем появлялись дицентрики, и вторичные перетяжки становились резко выраженными (Wolman и др., 1964).
В обоих рассмотренных исследованиях обращает внимание тот важный факт, что появление хромосомных аберраций предшествует или совпадает с процессом морфологической трансформации культуры. В полностью трансформированной гетероплоидной культуре практически каждая клетка содержит одну или больше аберрантных хромосом. Очевидно, эти клетки произошли от единичных клеток с ранними хроматидными разрывами, появившимися в культуре 6—7 пассажей ранее.
Таким образом, трансформацию культуры Moorhead и Saksela рассматривают как последствие изменений в наследственном материале клеток,, вызванных непосредственным или косвенным действием вируса.
А. Н. Мустафина (1966) наблюдала трансформацию диплоидных клеток различных культур через 2—3 месяца после их инфицирования. Она также обнаружила многочисленные нарушения в хромосомных наборах — хроматидные и хромосомные перестройки, эндоредупликации и полиплоидизацию. Однако этот автор полагает, что поражение ядерного аппарата клетки является не причиной, а следствием процесса трансформации.
Sticłi с соавторами (1964) использовали в исследовании действие вируса герпеса на культуру клеток человека и культуру нормальных диплоидных клеток хомячка. В культуре диплоидных клеток эмбрионального легкого1 человека через 24 часа до 67% делящихся клеток оказались содержащими поврежденные хромосомы. В клетках хомячка аберрации хромосом представляли собой хроматидные просветы и хроматидные разрывы; хромосом- гшх разрывов и обменов обнаружено не было.
Aula и Saksela (1966) исследовали действие вируса параинфлюэнцы (вирус Сендай) при инфицировании им культуры Hela и диплоидных культур клеток человека. На обоих типах культур обнаружено сходное изменение хромосом: окрашиваемость их снижается, хромосомы деспирализуются. Далее хроматиды в районе центромеры разъединяются. Благодаря дифференциальной деспирализации отдельных районов уплотненные участки хромосомы чередуются с нитевидными. В конечной стадии хромосомы претерпевают прогрессирующую фрагментацию, сходную с «пульверизацией», обнаруженной при исследовании действия на хромосомы вируса кори (Nichols, 1965). Обращает на себя внимание чрезвычайно ранняя реакция хромосом на действие вируса: первые их изменения обнаруживаются уже через 10 минут после инфицирования. Aula и Saksela полагают, что механизм повреждающего действия вируса параинфлюэнцы на хромосомы состоит в энзиматическом растворении ее участков.