Половой хроматин представляет собой морфологическое выражение феномена компенсации дозы генов. В экспериментах на культуре клеток было выяснено также, что поведение полового хроматина коррелирует с поведением в интерфазе поздно реплицирующейся Х-хромосомы.
Исследования полового хроматина, проводимые параллельно с исследованиями хромосом, продемонстрировали, что имеется тесное взаимоотношение между количеством и структурой Х-хромосом и количеством и размерами телец полового хроматина. В целом ряде работ специально разбирается вопрос об этом отношении (Grumbach, Morishima. 1962; Schwarzacher, 1962; Barr, Carr, 1962; Barr, 1964; 1966). Эти соотношения в общем сводятся к следующему.
1. У индивидуумов, имеющих одну Х-хромосому во всех клетках, эта хромосома деспирализована, подобно аутосомам, и находится в эухроматическом состоянии, благодаря чему в интерфазном ядре она морфологически не выявляется. Хроматинотрицательные клетки (клетки, ядра которых не содержат телец полового хроматина) обнаруживаются у индивидуумов, имеющих набор хромосом 45, ХО (синдром Тернера), 46, XY (нормальные мужчины), 47, XYY (синдром Клайнфельтера с двумя Y-хромосомами). В случае мозаицизма типа ХХ/ХО количество хроматинположительных ядер уменьшено против нормы.
2. У индивидуумов, имеющих две Х-хромосомы, в большинстве клеток одна хромосома неактивна, что морфологически выражается в ее готеропикнозе или гетерохроматизации; такая гетеропикнотическая хромосома в интерфазном ядре видна в виде тельца полового хроматина. Клетки, в которых ядра имеют одно тельце полового хроматина, обнаруживаются у индивидуумов, имеющих хромосомный набор 46, XX (нормальные женщины), 47, XXY (классический синдром Клайнфельтера), 48, XXYY (также синдром Клайнфельтера).
3. Сколько бы ни прибавлялось в хромосомном наборе Х-хромосом, только одна остается активной, эухроматизированной. Все лишние Х-хромосо- мы неактивны, гетерохроматнзированы и дают тельце полового хроматина. Два половых хроматина обнаруживаются у индивидуумов типа 47, XXX (синдром тришю-Х), 48, XXXY (синдром Клайнфельтера), 49, XXXYY (синдром Клайнфельтера). Три тельца полового хроматина обнаруживаются в ядрах клеток индивидуумов, имеющих хромосомный набор тина 48, ХХХХ (синдром, фенотипически мало отличающийся от синдрома трипло-Х), и у индивидуумов типа 49, XXXXY (более тяжелый клинически синдром Клайнфельтера).
4. При структурных аномалиях Х-хромосомы происходит изменение размеров полового хроматина. Маленькое тельце с уменьшенным содержанием ДНК (определенным цитофотометрически) обнаруживают в случае долеции Х-хромосомы. Увеличенное против нормы тельце с большим содержанием ДНК обнаруживают при наличии изохромосомы X но длинному плечу.
Подробное описание указанных аномалий дано в главе, посвященной заболеваниям, связанным с дефектами половых хромосом. Взаимоотношения, существующие между половым хроматином и половыми хромосомами, представлены на рис. 42.

Важным фактом является то, что в полиплоидных клетках число телец полового хроматина соответствует степени плоидности. Такого рода наблюдения принадлежат Klinger и Schwarzacher (1956, 1960), а также Hamden (1961).
Последний автор предложил формулу, позволяющую продемонстрировать зависимость количества телец полового хроматина от степени плоидности клеток. Согласно этой формуле, число телец полового хроматина равно числу Х-хромосом минус степень плоидности, деленную на два:
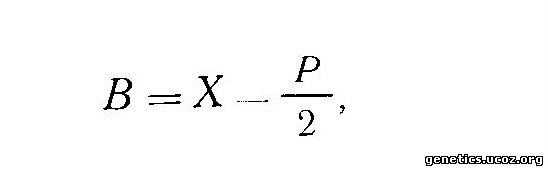
где В — число телец полового хроматина, X — число Х-хромосом, Р — степень плоидности.
Из этих наблюдений следует, что один диплоидный набор может содержать только одну гетерохроматизированную Х-хромосому. Что касается неполиплоидных клеток, то существует эмпирическое правило, согласно которому число телец полового хроматина равно числу Х-хромосом минус единица (В = Х—1). В анэуплоидных клетках (ракового происхождения или в длительной культуре) часто наблюдается снижение доли клеток, имеющих половой хроматин.
Это снижение обычно идет параллельно изменениям кариотипа и нарастанию анэуплоидии (James, 1960).
В 1954 г. Davidson и Smith (1954) обнаружили полоспецифические различия в ядрах полиморфноядерных лейкоцитов. Эта находка была подтверждена многими авторами и сводку этих данных можно найти в обзоре Davidson (1966). Большой вклад в классификацию полоспецифических образований полиморфноядерных лейкоцитов внес Kosenow (1956). Эти полоспецифические образования имеют вид характерных придатков, расположенных на лопастях ядер полинуклеаров. По виду их принято называть «барабанными палочками», или «drumstik» (рис. 43).
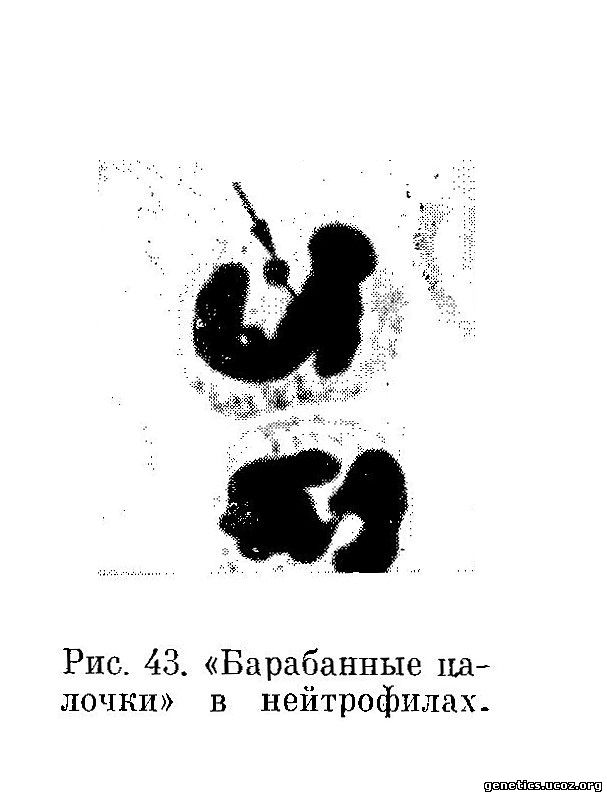
Они имеются в 1—6% нейтрофилов нормальных женщин и совсем не встречаются у мужчин. В указанных работах продемонстрировано, что «барабанные палочки» в нейтрофилах аналогичны половому хроматину или тельцам Барра в других клетках. Между полоспецифическими образованиями нейтрофилов и половыми хромосомами существуют такие же взаимоотношения, как между половым хроматином и половыми хромосомами (Fraccaro и др., 1964; Davidson, 1966). А. В. Капустігн (1965) наблюдал изменения в количестве барабанных палочек у женщин при беременности плодом мужского пола. Некоторые авторы (Greenblatt, Aydar, Gibson, 1962) отмечают, что в ряде случаев бывают расхождения между тем, что наблюдают в клетках слизистой рта и в нейтрофилах. Если исключить ошибки при микроскопировании, то это расхождение чаще всего можно объяснить мозаицизмом типа ХХ/ХО или XX/XY (Barr, Carr, 1962; Bain, Scott 1965) или случаями естественного или искусственного химеризма (Davidson, Fowler, Smith, 1958).
Половой диморфизм обнаружен также в малых плотноядерных лимфоцитах человека (В. А. Флоренсов, 1966).