Наши сведения за последние годы о строении и поведении хромосом в мейотической профазе в семенниках значительно обогатились. Хромосомы в стадии лептотены, зиготены и диплотены исследованы в семенниках субъектов от 18 лет до 81 года.
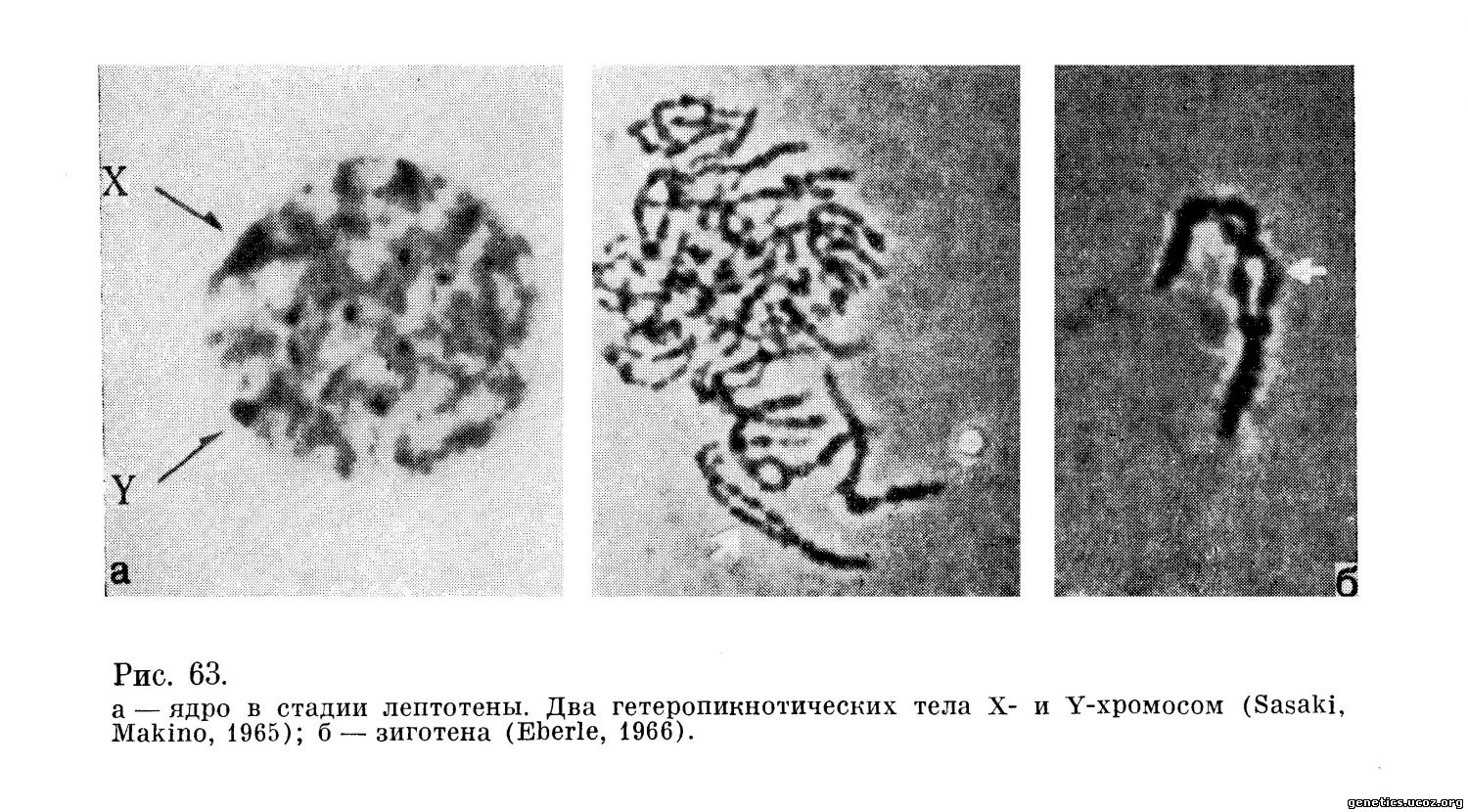
Лептогена. Ядра в стадии лептотены хорошо идентифицируются: они почти в 2 раза меньше по размеру, чем ядра в стадии пахитены или диплотены, и содержат очень тонкую, нежную сеть деспирализованных, слабо красящихся хромосом. Два отдельных гетеропикнотических тела неравной величины обычно располагаются у ядерной оболочки — это хромосомы X и Y (рис. 63, а).
Eberle (1966) показал, что многочисленные мелкие хромоцентры, обнаружимые в предмейотическом покоящемся ядре и в ядре ранней лептотены, в поздней лептотене полностью деспирализуются.
Зиготена. Стадия зиготены, когда гомологичные хромосомы находятся в процессе сдваивания, в сперматогенезе человека очень коротка, и данные о ней весьма ограничены (рис. 63, б). Конъюгация хромосом сопровождается их спирализацией, утолщением и быстрым переходом в стадию пахитены.
Пахитена. Эта стадия в сперматогенезе человека изучена достаточно полно (рис. 64). Бивалентные хромосомы спирализуются, укорачиваются и становятся обнаружимыми, как отдельные индивидуальные элементы. До момента поздней пахитены щель между гомологами в биваленте не обнаруживается.
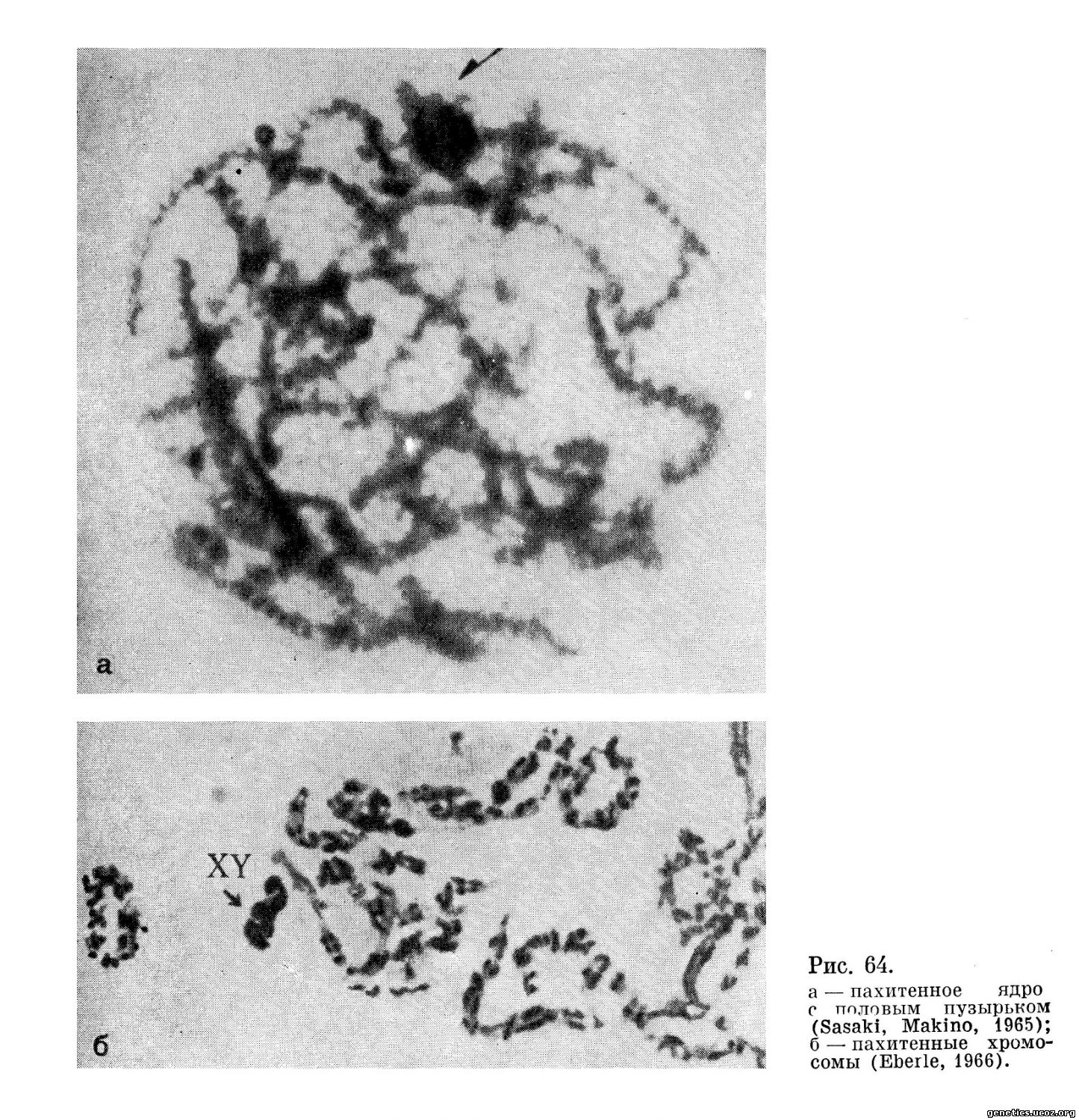
Уже в 1946 г. Schultz впервые на препаратах давленых кусочков семенников, окрашенных ацеторсеином, сделал попытку анализа пахитенных хромосом человека. Он установил, что отдельные хромосомы имеют определенный хромомерный рисунок и что гетерохроматические районы хромосом имеют тенденцию к общему соединению.
Позднее Yerganian (1957) исследовал изолированные пахитенные хромосомы и установил, что хромосомы в этой стадии имеют четкое хромомерное строение. Кроме хромомерного рисунка, характерного для индивидуальных хромосомных пар, каждый бивалент обладает отчетливыми, хорошо идентифицируемыми структурными особенностями: крупными гетерохроматическими узлами, сегментами с диффузным хроматином и, наконец, ядрышками, связанными с определенными участками некоторых хромосом.
Выяснение этих тонких структурных особенностей индивидуальных хромосом позволило составить пахитенные хромомерные карты для 9 бивалентных хромосом человека.
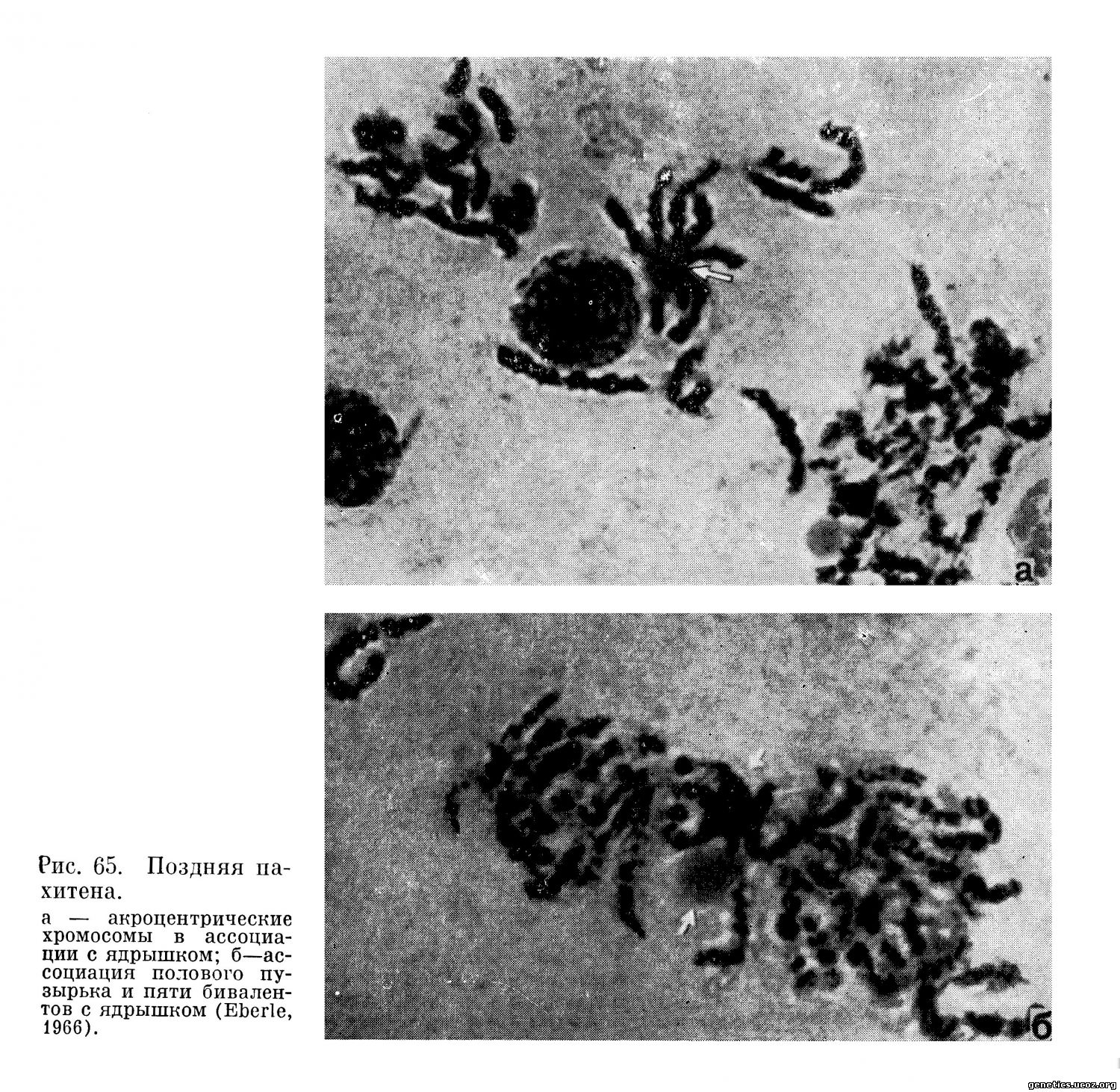
Наиболее обстоятельно преобразования хромосом на протяжении ранней, средней и поздней пахитенной стадии исследованы Sasaki и Makino (1965) и Eberle (1966). На основании тщательного анализа длины бивалента, размеров и расположения гетерохроматических сегментов Eberle дал структурные и количественные характеристики каждому из 22 аутосомных бивалентов в пахитене и представил их в виде кариограммы в соответствии с Денверской классификацией. Общая длина всех бивалентов в средней пахитене (без X- и Y-хромосом) равна 350,4 ц.
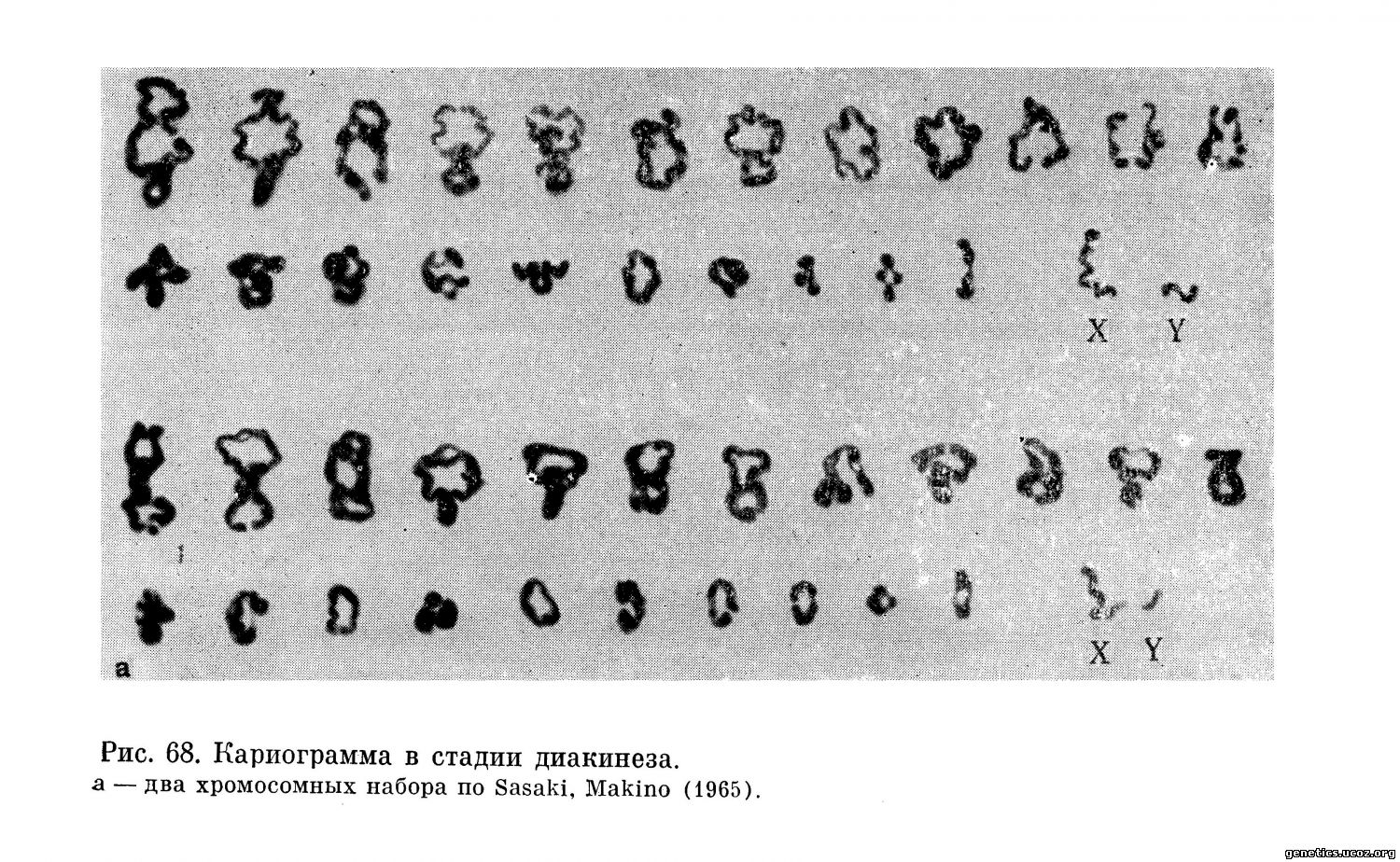
Согласно данным Eberle, в пахитене, диплотене и диакинезе биваленты идентифицируются не труднее, чем в метафазе митоза, что он продемонстрировал, представив разложенные по Денверской системе хромосомы в стадии диплонемы и диакинеза.
Как важнейшую особенность каждого бивалента Eberle рассматривает наличие в них гетеропикнотических сегментов (гетерохроматин). Согласно его данным, хромосомы 11, 12, 16 и 20 в классическом смысле полностью эухроматичны.
Хотя идентификация хромосом в трех кариограммах, составленных Eberle по стадиям мейоза, вряд ли может выдержать строгий критический анализ, его попытка весьма стимулирующая. Несомненно, дальнейшие исследования в этой области приведут к составлению полных и точных структурных и количественных характеристик индивидуальных хромосом человека в пахитене, диплонеме и диакинезе.
Составление карт хромосом в этих стадиях имеет огромное значение для выяснения характерных особенностей каждой из 23 пар хромосом человека.
В случае структурных изменений в хромосомном наборе, даже необнаружимых при анализе хромосом в митозе в соматических клетках, конъюгация гомологичных хромосом в мейозе нарушается.
Диплотена. Аутосомные биваленты в стадии диплотены представляют собой утолщенные нитеобразные элементы с хорошо выраженными хиазмами между гомологами (рис. 66).
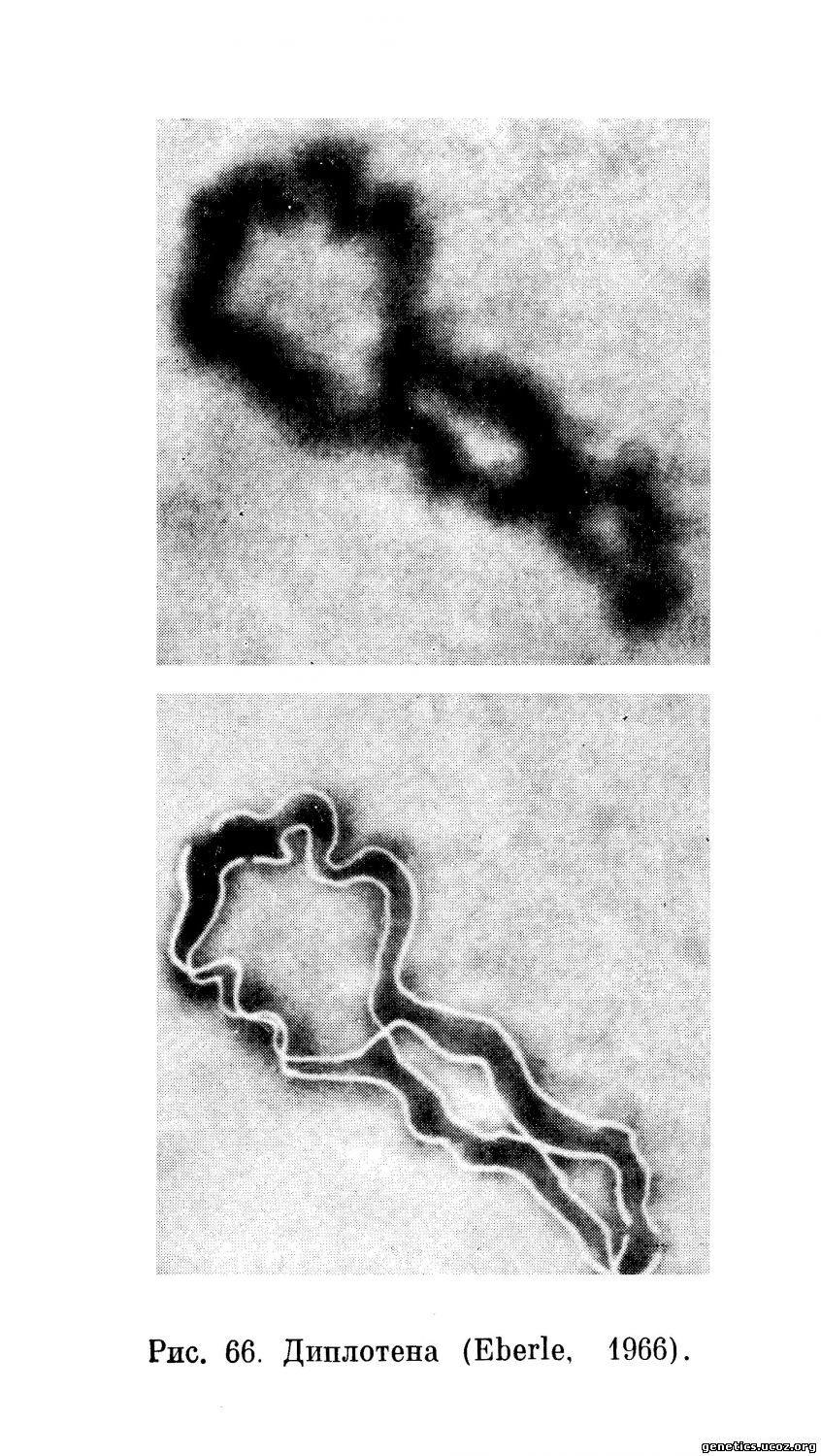
Многочисленность хиазм в диплотене сперматогенеза человека отмечается всеми исследователями, начиная с Roller (1937). В наиболее крупном биваленте Ford наблюдал до 5 хиазм (1956), Eberle — до 8 хиазм (1966). Среднее число хиазм на клетку у этих исследователей находится в хорошем соответствии: 55,9 по Ford и 55,7 по Eberle. Принимая условно, что каждая хиазма соответствует единичному кроссинговеру и что одна хиазма соответствует 50 сантиморганам, Ford определил общую генетическую длину хромосом человека в 27,9 морганид. Таким образом, его расчеты показали, что генетическая длина хромосом человека в 2 раза больше, чем у мыши, у которой она определена равной 19,2 морганиды (Slizinsky, 1955) или 16,2 морганиды (Carter, 1955). Ford весьма обоснованно полагает, что высокая частота хиазм представляет одну из важных причин, по которой у человека так редко обнаруживается аутосомное сцепление признаков.
Диакинез. Стадия диакинеза длится несколько дольше, чем диплотена. Прогрессивная спирализация хромосом сопровождается укорочением и утолщением бивалентов и повышением интенсивности их окрашиваемости. Отталкивание центромерных участков приводит к терминализации хиазм (рис. 67, 68). Их число на клетку от диплотены к позднему диакинезу снижается с 55,7 до 44,0.
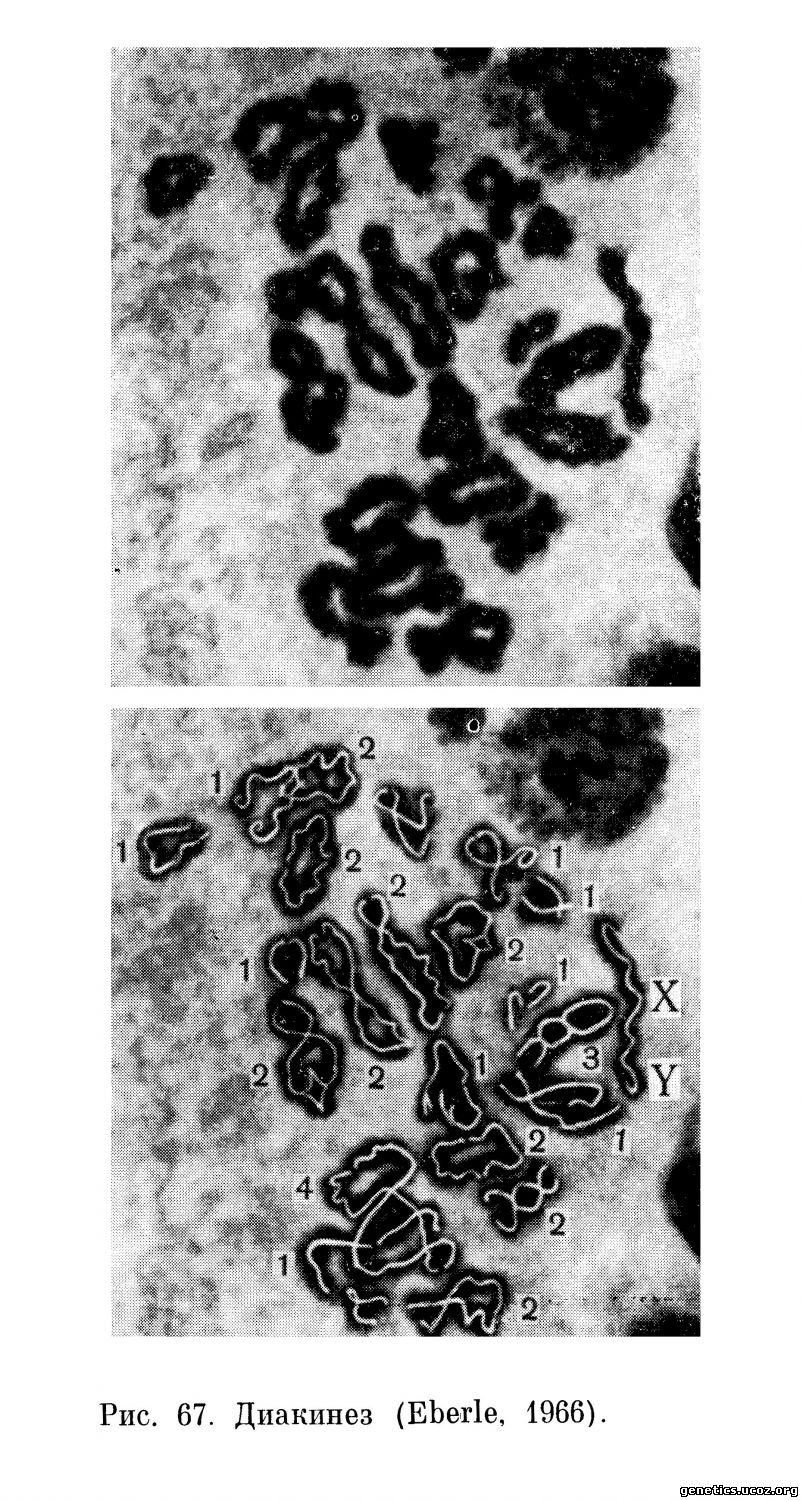
Систематизированные хромосомные наборы на стадии диакинеза представлены на рис. 68.

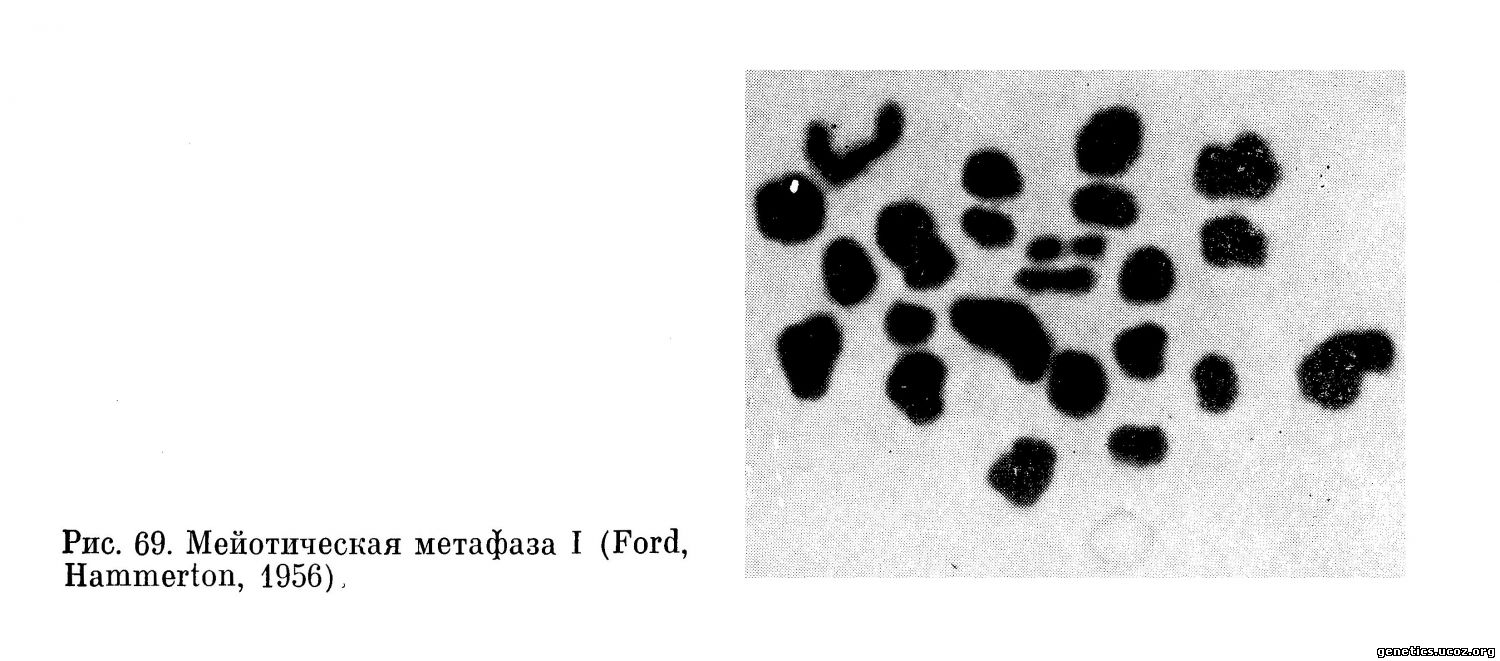
Первое мейотическое деление. При расположении бивалентов в экваториальной плоскости первого деления отчетливо видно строение каждого из двух гомологов, ориентированных центромерными районами к противоположным полюсам клетки.
Таким образом, закономерное расположение бивалентов на экваторе веретена с неизбежностью приводит к тому, что в первой мейотической анафазе к противоположным полюсам отходят целые гомологичные хромосомы, а не их хроматиды (рис. 69, 70).
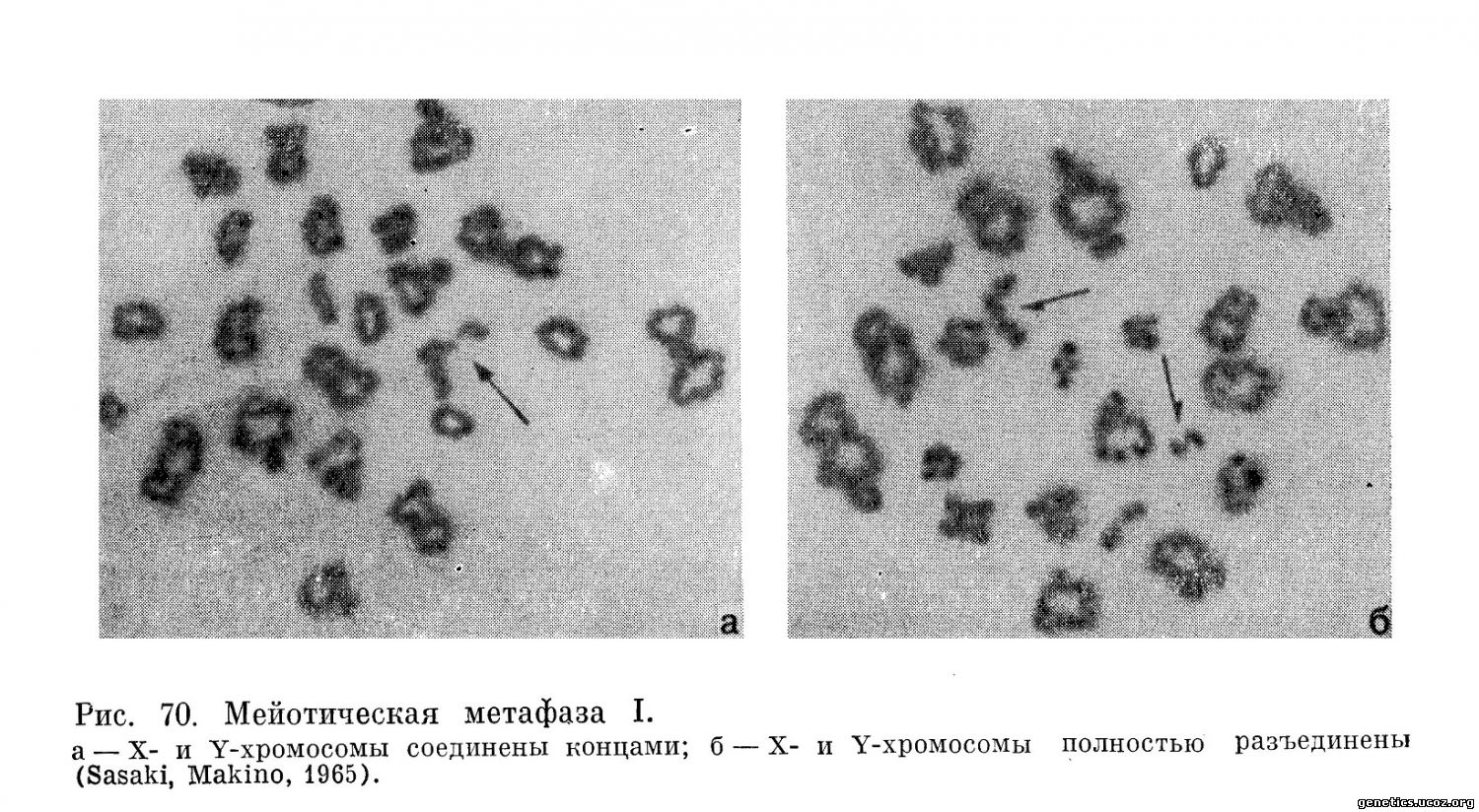
Первое мейотичеекое деление приводит к редукции числа хромосом. В телофазе I хромосомы, достигнув полюсов, слабо деспирализуются и переходят в состояние очень непродолжительного интеркинеза.