Гомологичные аутосомы одного и того же хромосомного набора очень часто отчетливо различаются между собой особенностями репликации. Это может выражаться в различиях общего количества метки или ее распределения по длине хромосомы или в том и другом. В какой степени эти различия обусловлены случайной вариацией в таких сложных процессах, как включение тимидина-Н3 в хромосомы и авторадиография, или закономерными различиями свойств хромосом отцовского и материнского происхождения — до настоящего времени не выяснено. Этот вопрос представляет собой сложную статистическую проблему.
Первые наблюдения об асинхронности репродукции гомологичных аутосом в конце периода синтеза в культуре лейкоцитов опубликовал Lima-de-Faria (1961). German (1962, 1964) и Bader с сотрудниками (1963) подтвердили эти наблюдения для хромосом той же культуры. Однако исследование английских цитогенетиков, использовавших обширную статистическую обработку материалов, не дало оснований для заключения об асинхронности репродукции гомологичных аутосом в культуре лейкоцитов периферической крови (Gilbert и др., 1962, 1965).
Обстоятельное исследование этого вопроса было проведено на первичной культуре эмбриональных фибробластов (А. А. Прокофьева-Бельговская и С. И. Слезингер, 1965). В этом исследовании проведен статистический анализ количества гранул серебра над гомологичными хромосомами 1-й и 2-й пары в начале и конце периода синтеза.
Установлено, что в начале периода синтеза эти различия недостоверны для хромосомы 1, но достоверны для хромосомы 2, вероятность нулевой гипотезы менее 5%. В конце периода S обнаружены значительные различия в интенсивности мечения как гомологов хромосомы 1, так и гомологов хромосомы 2 (вероятность нулевой гипотезы менее 0,1%) (рис. 55).
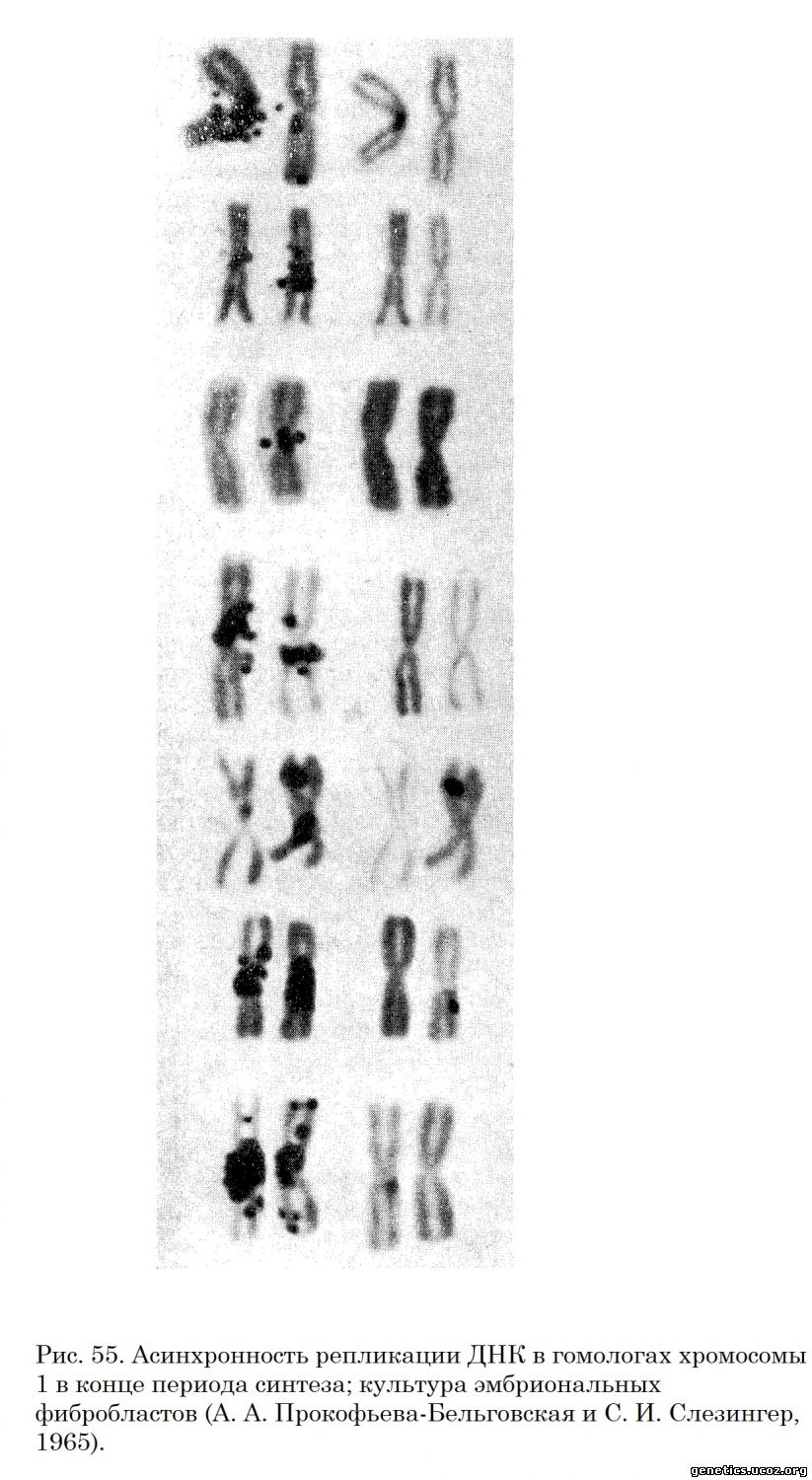
Анализ различия в репликации ДНК в гомологах хромосом 1 и 2 у человека показывает, что обнаруженная асинхронность в этом процессе может быть обусловлена двумя причинами:
1) разновременностью вступления участков гомологичных хромосом в период синтеза ДНК: при одинаковой скорости процесса репликации ДНК в обоих гомологах у одного из них период синтеза заканчивается раньше, чем у другого;
2) различиями скоростей процесса репликации ДНК; оба гомолога в этом случае вступают в период синтеза одновременно или разновременно.
Есть основания полагать, что гетероцикличность (или асинхронность) в системе гомологичных хромосом, если она подтвердится дальнейшими исследованиями, создается обоими механизмами.
Противоречивость заключений различных исследователей о степени синхронности репродукции гомологичных хромосом может быть объяснена рядом причин: различиями типов исследуемых клеток, возраста индивидуумов, наконец, методов статистической обработки результатов. Таким образом, проблема репродукции гомологичных аутосом требует дальнейшей разработки. Решение ее имеет большое значение для ряда вопросов, связанных с идентификацией хромосом в группах В, D и G на основании особенностей рисунка включения тимидина-Н3 в эти хромосомы.
Недавно обстоятельный анализ репликации ДНК в гомологичных хромосомах человека был проведен Сандбергом с коллегами (Sandberg et al. 1968). Они пришли к заключению, что для репликации гомологов некоторых пар хромосом (16, 17 и 18) весьма характерна антисинхронность — репликация ДНК одного локуса на этих хромосомах предохраняет от одновременной репликации ДНК в гомологичном локусе. Для гомологов всех хромосом, согласно их представлениям, характерна метасиихронность. Она характеризуется различием во времени репликации ДНК в гомологичных локусах (антисинхронность), причем этот промежуток времени может варьировать от локуса к локусу, от хромосомы к хромосоме и от одного к другому моменту периода S.