Структурные изменения хромосом у человека, хотя они и встречаются намного реже, чем численные аберрации, все же поставляют известную часть больных хромосомными заболеваниями. В общей генетической и цитогенетической литературе накоплен большой материал о происхождении, природе и поведении перестроенных хромосом. Так как в общих чертах и во многих деталях хромосомы высших животных и растений весьма сходны между собой, знание этого материала необходимо для исследования структурных перестроек собственно у человека.
Структурные перестройки можно классифицировать, исходя из нескольких принципов.
Во-первых, в момент возникновения они могут быть хромосомными или хроматидными. В том случае, если разрывы и соединения затронули только одну хроматиду из двух, перестройка возникает как хроматидная. Если же в момент возникновения хромосома состоит только из одной нити (т. е. в анафазе, телофазе и периоде G1 интерфазы), возникшая перестройка после первой же репликации становится хромосомной благодаря репродукции. В том случае, если перестройка произошла после редупликации и затронула одну хроматиду из двух, в первой после момента перестройки метафазе она будет выглядеть как хроматидная, в следующей же метафазе она также превратится в хромосомную. В том случае если перестройка произошла после репродукции и затронула обе хроматиды, она возникает как изохроматидная.
Во-вторых, структурные изменения хромосом могут сопровождаться изменением количества генетического материала (делеции и дупликации) или могут не сопровождаться изменением количества материала, а сводиться только к его перемещениям (инверсии, транслокации).
В-третьих, перестройки могут затронуть одну хромосому, обе хромосомы из пары гомологичных, две или больше негомологичные хромосомы.
В-четвертых, перестройки могут быть результатом одного или нескольких разрывов и соединений и, наконец, они могут быть способны или неспособны передаваться в сбалансированном состоянии. Последнее обстоятельство особенно важно для исследователя, работающего в области медицинской цитогенетики.
Сбалансированным по перестройке будет такой кариотип, в котором присутствуют все локусы всех входящих в кариотип хромосом, однако расположение их в пределах хромосомы или между хромосомами отличается от исходного нормального.
Несбалансированное состояние ранее сбалансированной перестройки выражается у ее носителя в делеции и (или) дупликации. Всякого рода перемещения участков хромосом в момент своего возникновения бывают сбалансированными, т. е. состоят только в изменении локализации одного или нескольких сегментов, но не в утрате или удвоении их в хромосомном наборе. Таким образом, несбалансированный кариотип может нести первичную делецию или дупликацию, или может быть результатом некоторых событий, имеющих место в мейозе у клетки, несущей сбалансированный кариотип. Поэтому мы вправе рассматривать отдельно первично и вторично несбалансированные кариотипы.
По типам различаются следующие перестройки.
1. Делеция, или нехватка. Это утрата части хромосомного материала, происходящая в результате двух разрывов и одного воссоединения с утратой сегмента, лежавшего между разрывами (рис. 18, а).
В приведенном случае один разрыв прошел между точками а и Ь, а другой — между точками с и d. Дистальные участки соединились, а ингерстициальный участок b—с утерялся. Организмы, гетерозиготные по делеции, гемизиготны по утраченным локусам, т. е. гены, локализованные в этом участке, будут представлены у него не парой аллелей, как в норме, а только одним аллелем. В результате этого все рецессивные гены, в том числе и летальные, лежащие в соответствующем месте интактного гомолога, проявляются фенотипически.
У человека описаны делеции по хромосомам 18-й пары, по хромосомам группы В и некоторым другим. Делеция по одной из хромосом группы В, впервые обнаруженная Lejeune и Turpin, клинически выражается в синдроме так называемого кошачьего крика.
Крайне редко встречается особый тип делеции, так называемая «концевая нехватка», когда в результате одного единственного разрыва часть хромосомы, не несущая центромеры (ацентрический фрагмент), теряется из ядра и резорбируется в цитоплазме. Хотя концевая нехватка происходит в результате единственного разрыва, она встречается редко, так как в результате концевой нехватки теряется теломера — жизненно важный структурный компонент хромосомы. Если же в случае «концевой нехватки» разрыв проходит через саму теломеру, то часть ее остается. Хромосома без теломеры, как правило, неспособна к дальнейшему существованию. Это не относится к так называемым кольцевым хромосомам, возникающим в результате двух концевых нехваток с последующим соединением разорванных проксимальных концов (рис. 18, б).
К делеции (и дупликации) относится еще один тип перестройки —образование изохромосомы, которое будет рассмотрено отдельно.
Делецию впервые открыл Bridges на Drosophila melanogaster в 1917 г. Одна самка первого поколения из скрещивания Ваг не наследовала доминантный сцепленный с полом ген Ваг. В потомстве этой самки наблюдалось необычное соотношение полов 2:1. Это указывало, что половина мужских зигот гибнет, не доходя до стадии имаго. Этими погибшими самцами были те, которые получили хромосому X с делецией участка, несущего ген Ваг.

2. Дупликация. Это удвоение сегмента хромосомы, в результате которого клетка или организм, несущий дупликацию, становится гиперплоидным по данному сегменту. Первично дупликация может возникнуть в результате неравного кроссинговера. Неравный кроссинговер имеет место в случае, если в некотором участке хромосомы гомологичные локусы не конъюгируют в профазе I мейоза точно, а сдвигаются один относительно другого на некоторое расстояние (рис. 19).
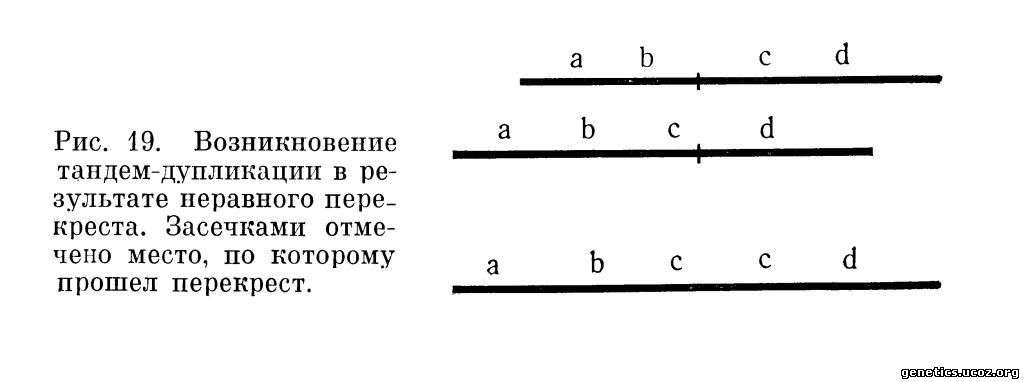
Разрешающая способность методов анализа хромосом человека не позволяет точно идентифицировать дупликацию, возникающую таким образом.
Все другие дупликации, за исключением изохромосомы, имеют не первичное происхождение.
Дуплицированный участок может быть расположен в исходной хромосоме; в этом случае он либо может лежать непосредственно за исходным участком (тандем-дупликация), либо быть в том же плече, что и исходный участок, или в другом плече исходной хромосомы. Кроме того, дуплицированный участок может быть локализован в негомологичной хромосоме.
Фенотипически дупликации менее неблагоприятны, чем делеции. Дупликация также была впервые обнаружена с помощью генетического метода Bridges на Drosophila melanogaster в 1919 г. Впоследствии (в 1935 г.) Bridges подтвердил это прямым наблюдением на гигантских хромосомах слюнных желез дрозофилы.
3. Изохромосомы. В результате разрыва хромосомы в районе центромеры возникают две акроцентрические хромосомы с концевым положением центромеры. Такие хромосомы нестабильны: их хроматиды, отталкиваясь друг от друга, формируют метацентрическую хромосому, у которой таким образом оба плеча, будучи сестринскими хроматидами, полностью идентичны и соединены центромерой. Так как каждая изохромосома состоит из двух совершенно идентичных плеч, она тем самым является дупликантной по этому плечу, и так как она утратила второе свое плечо, то имеет делецию по второму плечу. То же справедливо и для второй изохромосомы (рис. 20).

Если после анафазы две образовавшиеся изохромосомы попадут в разные дочерние ядра, то и эти ядра будут иметь делецию по одному плечу и дупликацию по другому. Поэтому поперечное деление центромеры приводит и к делеции, и к дупликации. Образование изохромосомы — единственная хромосомная аберрация, возникающая в результате одного разрыва.
Такие события нередко обнаруживаются у человека. Описаны изохромосомы X, Y и в группе 21—22. Появление таких хромосом всегда сопровождается соответствующими дефектами в развитии.
Механизм образования изохромосом был предложен Lima-de-Faria (1959) вместе с описанием строения и функционирования центромерного района.
Рассмотренные типы хромосомных перестроек представляют собой либо делецию, либо дупликацию (или то и другое вместе) уже в момент своего возникновения. Таким образом, они никогда не бывают сбалансированными. И как следствие этого фенотип их носителей всегда (или почти всегда исключения) отличается от нормы. Большинство таких перестроек вообще летально для их носителей, а те из носителей, которые выживают, как правило, неспособны оставить потомство. Для медико-цитогенетической консультации гораздо больший интерес представляют перестройки, которые в момент своего возникновения не приводят ни к утрате, ни к удвоению хромосомного материала, т. е. сбалансированные перестройки. Они способны передаваться в сбалансированном состоянии (за исключением перестройки одного типа) на протяжении трех и более (возможно, значительно больше) поколений, приводя время от времени к появлению в семье аномального потомства.
Поэтому совершенно очевидна необходимость анализа возможных последствий, возникающих в мейозе, в результате расхождения перестроенных хромосом из первично сбалансированного кариотипа. Хотя количественное соотношение тех или иных гамет, продуцируемых носителем сбалансированной транслокации, зависит от многих факторов, часть из которых еще не поддается учету, такой анализ позволяет заранее предусмотреть все возможные классы сбалансированных и несбалансированных гамет.
4. Инверсия. В случае инверсии участок хромосомы после двух разрывов поворачивается на 180° и разорванные концы соединяются в новом порядке. Если в инвертированный участок попадает центромера хромосомы, такую инверсию называют перицентрической, если же инверсия затрагивает только одно плечо хромосомы, она называется парацентрической. В метафазных хромосомах соматических клеток инверсию можно обнаружить только в тех случаях, когда разрывы проходят по обе стороны от центромеры (т. е. перицентрически) и на разных от нее расстояниях, так как в этом случае изменяется морфология хромосомы, а именно ее центромерный индекс, и хромосома может, например, из акроцентрической превратиться в метацентрическую.
Инверсии, так же как и другие виды хромосомных перестроек, впервые были описаны с помощью генетических методов. В 1931 г. Стертевант, сравнивая два вида Drosophila — Dr. melanogaster и Dr. simulans — убедился, что три идентичных (это было проверено тестом на аллелизм) рецессивных гена лежат у этих видов в разной последовательности, а именно: у Dr. melanogaster scarlet peach delta, тогда как у Dr. simulans scarlet deltoid peach.
Таким образом, гены в инвертированном участке хромосомы лежат в обратном по отношению к исходной хромосоме порядке. Сама по себе инверсия не обязательно изменяет фенотип носителя инвертированной хромосомы, однако перекрест внутри инвертированного сегмента приводит к крупным перестройкам. Рассмотрим их. Пусть в некоторой хромосоме имеется порядок генов А, В, С, D, E, F, G, Н (С обозначает центромеру). Рассмотрим вначале, что произойдет с такой хромосомой в мейозе, если в ней случится парацентрическая инверсия, т. е. не затрагивающая центромеры и касающаяся только одного плеча (рис. 21, а). В результате парацентрической инверсии возникнет новый порядок генов. Из приведенной схемы видно, что разрывы прошли между локусами D и Е (первый) и локусами G и Н (второй). В мейозе происходит точная, локус к локусу, конъюгация гомологичных хромосом. Для такой конъюгации хромосом придется образовать так называемую инверсионную петлю (рис. 21, б). Если в пределах инвертированного сегмента произойдет перекрест, то в результате возникнут две перестроенные хромосомы с новым порядком генов (рис. 21, в).
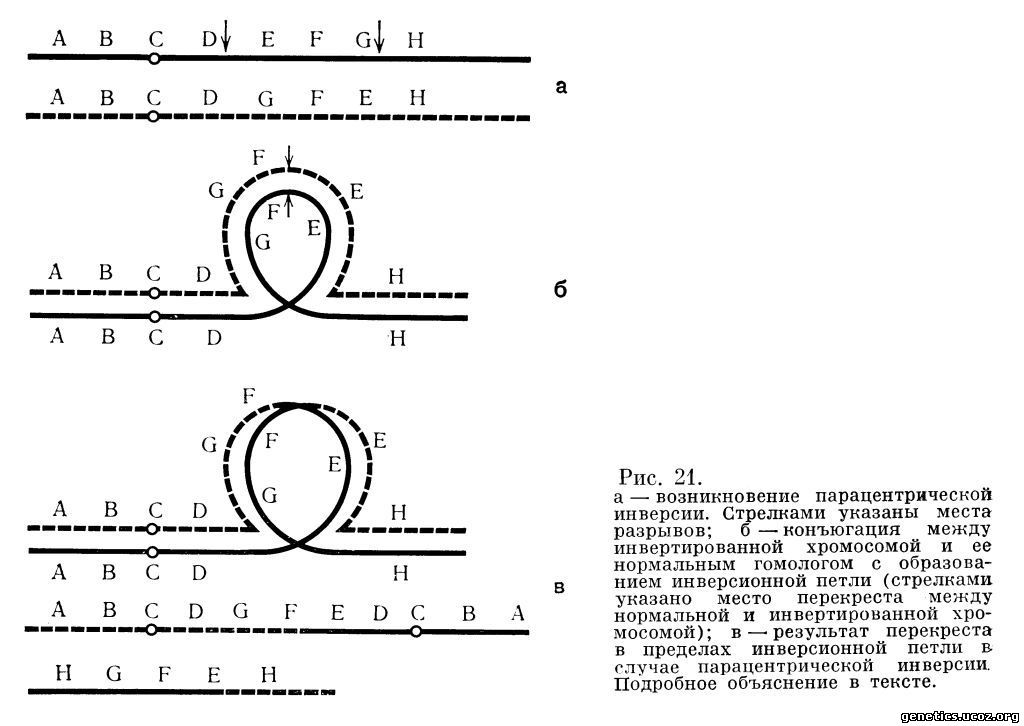
Как видно из рис. 21, и первый, и второй продукты перекреста имеют дупликацию одних участков и делецию других. Однако, кроме этого, у одного из них дважды представлена центромера. Такие хромосомы называются дицентрическими, или просто дицентриками. В том случае, если центромеры одной дицентрической хромосомы пойдут к противоположным полюсам, в анафазе I мейоза возникнет хроматидный мост, который в дальнейшем разорвется. Ясно, что зигота, получившая такую перестройку, будет нежизнеспособна. Второй продукт перестройки, помимо дупликации и делеции некоторых сегментов, совсем не имеет центромеры, что поведет к его утрате после анафазы I. Конечный результат будет тот же.
Таким образом, организм, гетерозиготный по парацентрической инверсии, будет давать то или иное количество гамет, приводящих потомство к гибели в стадии зиготы или несколько позднее.
Рассмотрим, что произойдет в случае перицентрической инверсии. Пусть разрывы процдут между генами А и В в одном плече и между генами F и G в другом. Тогда возникнет хромосома с новой последовательностью генов (рис. 22, а): А, F, E, D, С, В, G, Н.
Сразу же становится ясно, что в результате такой перестройки изменился центромерный индекс хромосомы, т. е. отношение короткого плеча ко всей длине хромосомы. В результате перекреста в инвертированном сегменте в этом случае возникнут две хромосомы с такой последовательностью генов в каждой из них: А, В, С, D, E, F, А и H, G, F, E, D, С, В, G, Н (перекрест в разбираемом случае произошел между генами В и С) (рис. 22, б, в).
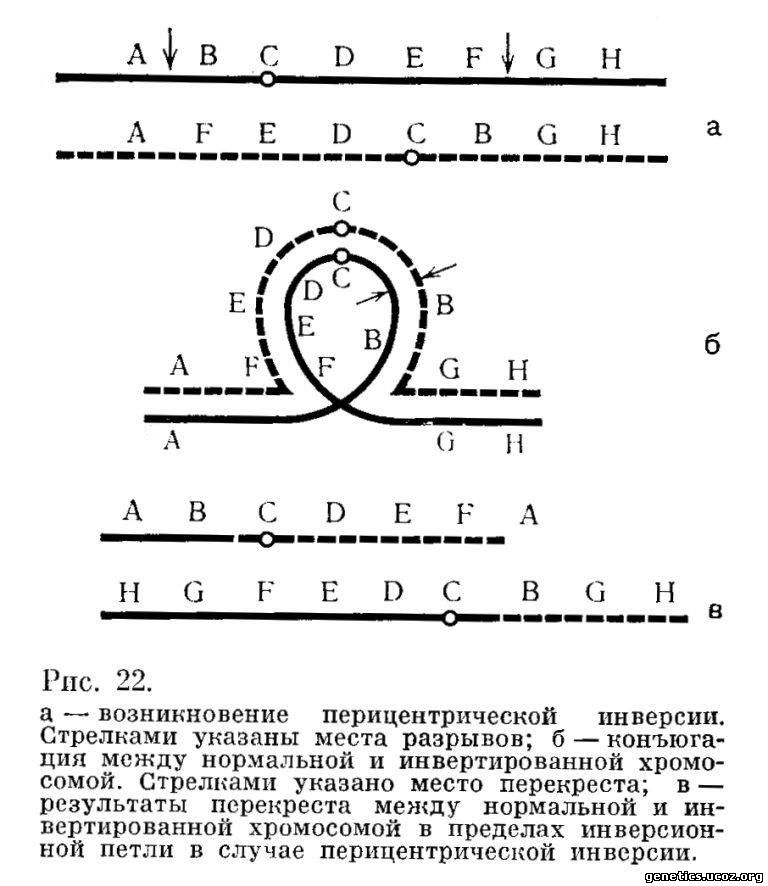
Обе возникшие в результате перекреста хромосомы имеют центромеру и способны к дальнейшему существованию. Однако нетрудно видеть, что обе они несут делецию и дупликацию. Первая имеет дупликацию района А и делецию района G и Н, вторая, наоборот, — делецию района А и дупликацию районов G и Н. Будут ли организмы, получившие такие хромосомы, жизнеспособны, зависит от размеров и роли этих участков. У человека, во всяком случае, такие примеры описаны.
5. Транслокации. Это обмен сегментами между хромосомами. Различают следующие типы транслокаций:
а) реципрокные, когда две хромосомы взаимно обмениваются сегментами;
б) нереципрокные, когда сегменты одной хромосомы переносятся в другую;
в) транслокации типа центрического соединения, когда после разрывов в околоцентромерных районах соединяются два несущих центромеры фрагмента таким образом, что две их центромеры соединяются, образуя одну.
а) Реципрокная транслокация
Рассмотрим механизм образования реципрокной транслокации. Пусть имеются две негомологичные хромосомы А и В. Если после разрывов, на¬пример в районах А5 и В5, участок A5-7 присоединится к участку В5, а участок В6-7 — соответственно к участку А4, то возникнет реципрокная транслокация (рис. 23).

Как видно из рис. 23, в результате реципрокной транслокации хромосомный материал из ядра не теряется, происходит только его транспозиция; таким образом, после реципрокной транслокации возникает сбалансированный кариотип. Кроме того, из схемы видно, что образовавшиеся после перестройки хромосомы отличаются по морфологии от исходных и, следовательно, перестройка доступна для микроскопического обнаружения. Нетрудно видеть, что это справедливо только для тех случаев, когда транслоцированные сегменты отличаются друг от друга по размерам.
В результате конъюгации в мейозе транслоцированные хромосомы вместе со своими неперестроенными гомологами образуют характерную для реципрокной транслокации фигуру так называемого «транслокационного креста» (рис. 24).
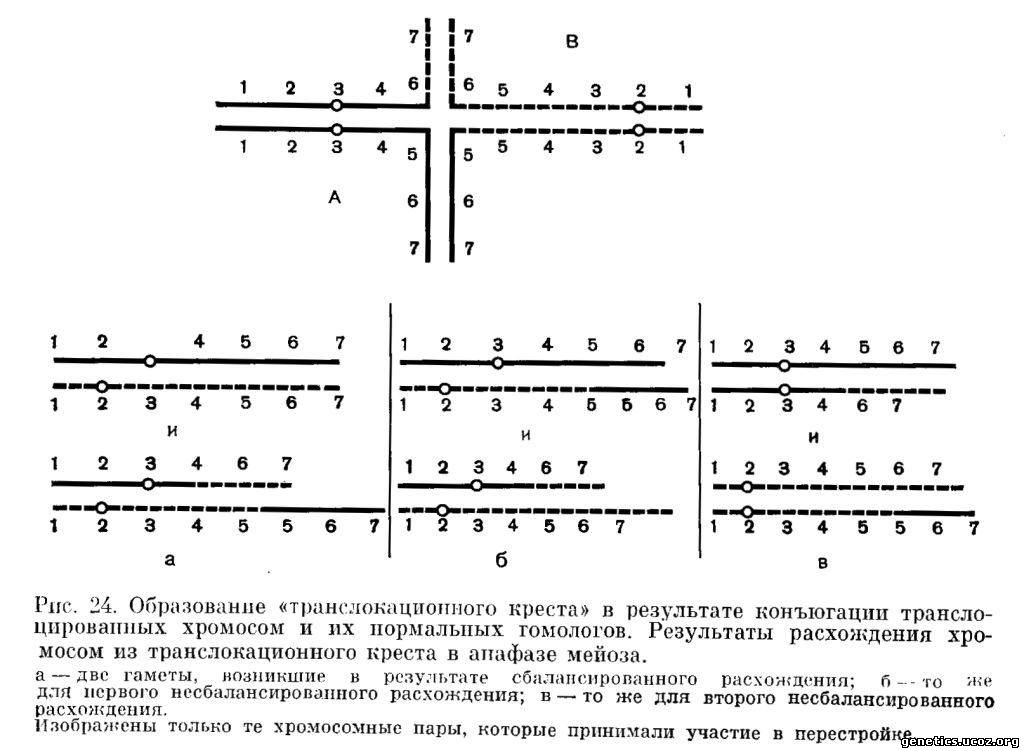
Так как в транслокациопном кресте конъюгируют 4 хромосомы, из них образуется не два бивалента, как в обычном случае, а одна сложная фигура — тетравалент. Поэтому морфологически различимых образований в метафазе I мейоза в клетке, несущей сбалансированную транслокацию, будет на одно меньше, чем в нормальной клетке.
В анафазе I мейоза возможны 7 типов расхождения хромосом из транслокационного креста, в результате которых возникнет 14 классов гамет.
1. В том случае, если неперестроенные хромосомы отойдут к одному полюсу веретена деления, а их перестроенные гомологи — к другому, возникнут гаметы двух типов. В одной гамете будут содержаться нормальные гомологи перестроенных хромосом, а в другой — сами перестроенные хромосомы. В результате такого расхождения поэтому каждый раз возникает одна гамета, несущая нормальный набор, и одна гамета, несущая перестроенный набор, но в сбалансированном состоянии, так как в ней присутствуют все участки всех хромосом, хотя и иначе друг относительно друга расположенные (рис. 24). Из таких гамет после оплодотворения разовьются фенотипически нормальные организмы, однако один из них, так же как и его родитель, будет обладателем сбалансированного по перестройке кариотипа. Такой тип расхождения мы будем называть в дальнейшем сбалансированным (рис. 24, а).
2. Если центромеры гомологичных хромосом оттолкнутся и отойдут к противоположным полюсам таким образом, что к каждому полюсу пойдет одна нормальная и одна перестроенная хромосома, то возникнут две гаметы, несущие несбалансированный набор хромосом. В каждой гамете один участок будет представлен дважды (дупликация), а другой будет отсутствовать (делеция) (рис. 24, б). Из рис. 24 видно, что в одной гамете участок A5-7 отсутствует, а участок В6-7 представлен дважды. В реципрокной гамете будет обратная картина, а именно участок А5-7 будет представлен дважды, а участок В6-7 будет отсутствовать. Таким образом, в результате подобного расхождения возникнут две гаметы, несущие несбалансированные хромосомные наборы. Мы будем называть такой тип расхождения I несбалансированным.
3. Если две гомологичные хромосомы (правильнее говорить о «гомологичных центромерах») отойдут к одному полюсу, а две другие гомологичные — к другому, то возникнут также два несбалансированных хромосомных набора. В каждой гамете, как это видно из рис. 24, в, неперестроенное плечо, центромерный район и сегменты, лежащие между центромерой и местом разрыва одного участника перестройки, представлены дважды, а те же районы другого участника перестройки не представлены совсем. Расхождение такого типа мы будем называть II несбалансированным. В результате II несбалансированного расхождения также возникают две гаметы, каждая из которых песет делецию одних участков и дупликацию других.
Остальные четыре типа расхождения хромосом из транслокационного креста состоят в том, что к одному полюсу веретена деления идет одна хромосома, а к другому — три остальные. Нетрудно видеть, что возможны именно четыре типа такого рода расхождения и что каждый раз будут возникать две гаметы, одна из которых несет только делецию по некоторым сегментам принимающих участие в перестройке хромосом, а другая, реципрокная ей, — дупликацию по том же участкам. В дальнейшем мы будем называть этот тип неравным расхождением. Таким образом, в результате подобного рода расхождений возникают анэуплоидные гаметы, половина из которых песет избыток, а другая половина — недостаток хромосомного материала.
Анализируя семьи, в которых встречается более одного случая хромосомной аномалии, исследователи часто обнаруживают там именно реципрокную транслокацию. Клиническая картина больных при этом, как правило, включает в себя обычную симптоматику хромосомных болезней, по каждая отдельная перестройка имеет и свои особенные симптомы .