Нереципрокная транслокация
Если участок одной хромосомы переносится в другую, негомологичную первой, хромосому, то такую перестройку называют не реципрокной транслокацией. Для осуществления нереципрокной транслокации необходимо наличие трех разрывов, за которыми следуют три соединения разорванных концов в отличном от первоначального порядке (рис. 25, а).
Пусть в хромосоме А произойдет разрыв между участками А2 и А3, а в хромосоме В произойдут два разрыва: первый между участками В3 и В4, а второй — между участками В5 и В6. Пусть участок В3 соединится с участком В6, а сегмент В4-5 попадет между сегментами А2 и А3. Такая перестройка и будет нереципрокной транслокацией. Так же как и в случае реципрокной транслокации, в этом случае не происходит ни утраты хромосомного материала из перестроенной клетки, ни его прибавления, т. е. в момент своего возникновения перестройка сбалансирована. Так как для возникновения нереципрокной транслокации необходимо образование трех разрывов, тогда как для возникновения реципрокной достаточно только двух, то естественно, что первые встречаются реже, чем вторые.
До недавнего времени нереципрокным транелокациям не уделялось внимания, однако сейчас они стали интересовать исследователей, работающих в области цитогенетики человека. В ряде случаев в результате анализа карпотипа больных, подозреваемых на наличие хромосомной перестройки, были получены данные о том, что у пораженного (чаще всего у пробанда) и у одного из его фенотипически нормальных родителей имеется морфологически неотличимый, но перестроенный кариотип. В этой связи высказывались различные соображения относительно причины характерной симптоматики у пробанда.
Однако в 1965 г. Lejeunc и Berger (1965) предложили объяснение этим наблюдениям, исходя из предположения о наличии в этих случаях нереципрокной транслокации. К настоящему времени опуоликовано еще несколько таких случаев.
Рассуждения Lejeune и Berger сводятся к следующему. Пусть между двумя пегомологичными хромосомами произошла нереципрокная транслокация (см. рис. 25, а). Для точной конъюгации, гомологичный локус к гомологичному локусу, перестроенным хромосомам придется образовать фигуру, несколько напоминающую инверсионную петлю (рис. 25, б). В образовании петли в этом случае принимает участие нормальный гомолог хромосомы-донора и сегмент хромосомы-донора, перенесенный в хромосому-реципиент. Хромосома-донор и нормальный гомолог реципиента конъюгируют с теми участками своих гомологов, которые собственно в перестройке участия не принимают.
Если в пределах конъюгационной петли не произойдет перекреста, то в результате расхождения хромосом в анафазе I мейоза теоретически могут возникнуть, в зависимости от типа расхождения, те же 14 классов гамет. Мы рассмотрим только сбалансированное и I несбалансированное расхождение, т. е. те случаи, когда гомологичные центромеры отталкиваются и расходятся к противоположным полюсам.
В случае сбалансированного расхождения в одну гамету попадут нормальный гомолог хромосомы-донора и нормальный гомолог хромосомы- реципиента (следует помнить, что донор и реципиент — негомологичные хромосомы), а во вторую гамету — хромосома-донор и хромосома-реципиент. В результате слияния первой гаметы с гаметой другого родителя возникнет нормальная особь с нормальным кариотипом, в результате слияния второй гаметы с гаметой другого родителя — фенотипически нормальная особь, несущая сбалансированную транслокацию, т. е. возникнет кариотип, аналогичный родительскому.
В случае I несбалансированного расхождения возникнут две гаметы; обе они будут нести несбалансированный набор хромосом. В одной из них будет делецпя, а в другой — дупликация перенесенного участка В4-5.
Гораздо больший интерес представляет рассмотрение тех случаев, когда в продолах конъюгационной петли произойдет один или любое нечетное количество перекрестов. (Четное количество перекрестов восстанавливает исходный порядок локусов справа и слева от области перекрестов; перекресты вне пределов конъюгационной петли не изменят рассматриваемую картину). Как следует из схемы (рис. 26). в перекресте принимают участие хромосома-реципиент и нормальный гомолог донора.
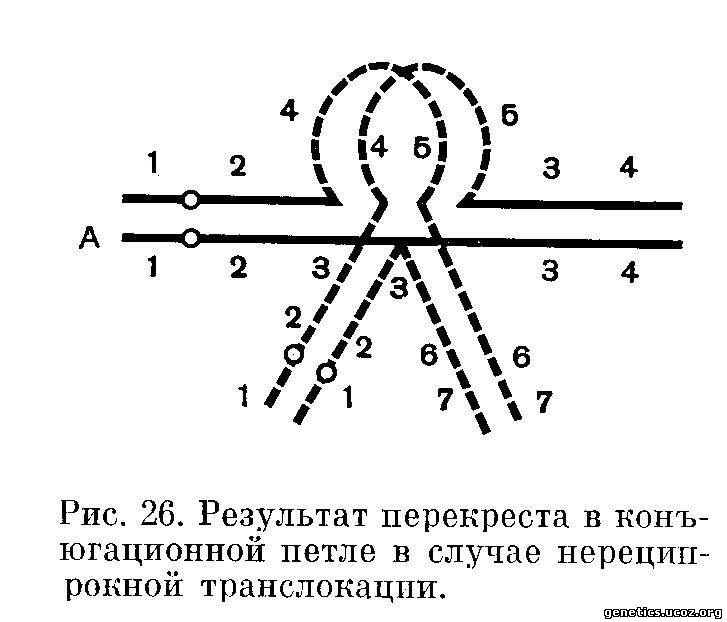
Пусть в пределах конъюгационной петли произойдет перекрест между локусами В4 и В5. В результате перекреста возникнут две хромосомы с новым, не существовавшим ранее набором локусов, а именно: хромосома-реципиент А (обозначается по принадлежности центромерпого района) будет состоять из локусов А1,2, В4,5,6,7, хромосома — нормальный гомолог донора В будет состоять из локусов В1,2,3,4,5, А3,4.
Таким образом, из 4 хромосом (две возникли в результате перекреста и две существовали ранее — нормальный гомолог реципиента и хромосома-донор) нот пи двух полностью гомологичных, ни двух полностью комплементарных, как это и видно из рис. 26. Поэтому пи один тип расхождения хромосом в анафазе I мейоза не даст ни нормальных, ни сбалансированных гамет. Рассмотрим один тип расхождения, который, по-видимому, имеет место наиболее часто, хотя окончательно судить об этом еще нельзя. Наиболее вероятным будет такой тип расхождения, в котором, во-первых, к противоположным полюсам разойдутся рекомбинантные хромосомы и, во-вторых, центромеры гомологичных хромосом также оттолкнутся друг от друга и разойдутся к противоположным полюсам. В нормальном течении мейоза обычно выполняются оба эти условия.
При соблюдении этих условий к одному полюсу отойдет возникшая в результате рекомбинации хромосома А (обозначение по-прежнему по центромере) с порядком локусов А1,2, В4,5,6,7 и хромосома-донор, т. е. В1,2,3,6,7; к другому полюсу отойдут хромосома В с порядком локусов В1,2,3,4,5, А3,4 и нормальный гомолог реципиента, т. е. А1,2,3,4 . Нетрудно видеть, что в первой гамете совсем не представлены локусы А3,4 и дважды представлены В6,7, а во второй — обратная картина, т. е. дважды представлены локусы А3,4 и совсем не представлены В6, 7. Таким образом, обе гаметы имеют делецию одних локусов и дупликацию других (рис. 27). Поэтому, если такие гаметы и дадут начало жизнеспособным особям, то последние, несомненно, будут фенотипически ненормальными.

В приведенной схеме перестройки (см. рис. 25, а) отрезки А3.4 и B6,7 равны по размеру, т. е. разрывы и в хромосоме А, и в хромосоме В (нижний) прошли па равном расстоянии от теломеры подвергшегося перестройке плеча. Это, разумеется, частный случай, — разрывы могут проходить на любом расстоянии от теломеры. Однако этот частный случай интересен тем, что после перекреста в конъюгационной петле возникают хромосомы, морфологически (т. е. по длине и центромерному индексу) идентичные исходным. Действительно, хромосома А после рекомбинации имеет 6 сегментов; столько же, сколько их было у хромосомы-реципиента, хотя состав локусов отличается от исходного. То же справедливо для хромосомы В, которая после рекомбинации состоит из 7 сегментов, так же как и нормальный гомолог донора. После слияния нормальной гаметы с гаметой, несущей рекомбинантную хромосому А и хромосому-донора (первая гамета на нашей схеме), возникнет несбалансированный (делеция А3,4 и дупликация В6,7), но морфологически неотличимый от сбалансированного родительского кариотип.
Не вдаваясь в дальнейшее рассмотрение всех возможных вариантов, отметим следующее: если отрезки между разрывом и теломерой у двух перестроенных хромосом неравны, то кариотип пораженного потомка будет отличаться от сбалансированного родительского, что и было показано авторами изложенной здесь теории, которую они предложили назвать «рекомбинационной анэусомией».
Во всех приведенных выше случаях мы рассматривали транслокации между негомологичными хромосомами. Однако в такую перестройку могут вступить и гомологи. Так как транслокация между гомологичными хромосомами, подобно другим транслокациям, в момент своего возникновения не приводит ни к добавлению, ни к утрате хромосомного материала из ядра, то в результате такой транслокации возникает клетка со сбалансированным хромосомным набором. Разумеется, транслокация между гомологичными хромосомами не может быть абсолютно реципрокной, так как обмен гомологичными сегментами с одними и теми же локусами будет уже но транслокацией, а кроссинговером. Поэтому независимо от того, является ли обмен взаимным или хромосома-донор не выступает одномоментно в качестве хромосомы-реципиента, все равно после транслокации между гомологичными хромосомами одна из них несет делецию некоторых локусов, а другая — дупликацию тех же локусов (рис. 28).
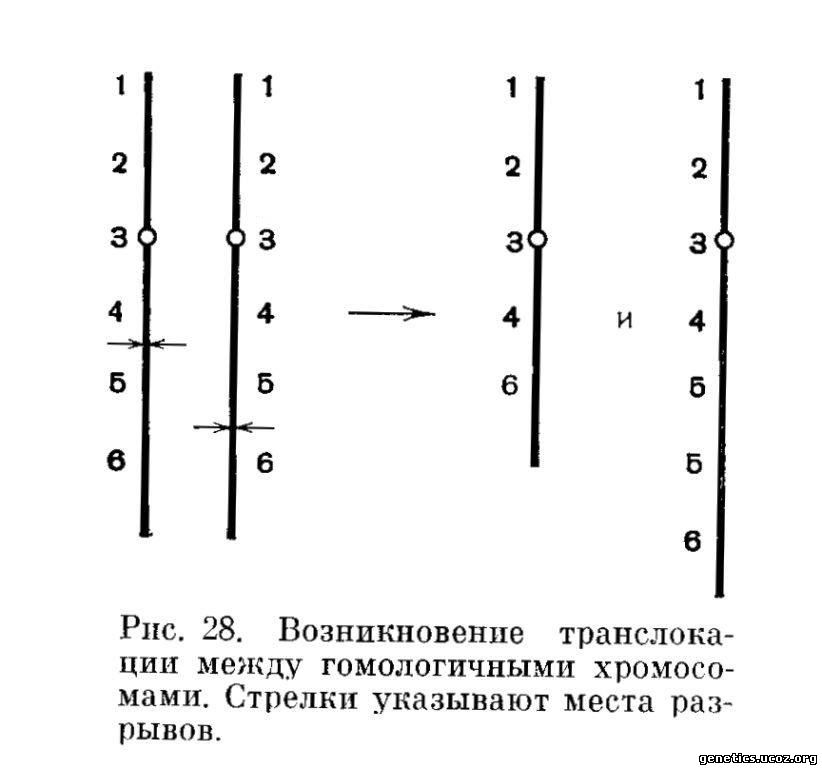
Так как в результате транслокации между гомологами обе вступившие в перестройку хромосомы становятся аберрантными, в рассматриваемом ядро отсутствуют нормальные хромосомы данной пары. Поэтому из такой клетки ни при каких обстоятельствах не может возникнуть в мейозе эуплоидных гамет, нормальных или сбалансированных. Носитель сбалансированной транслокации между гомологичными хромосомами является ее первым и последним обладателем, а у его потомков будет только делеция или дупликация по перестроенному сегменту.
Центрическое соединение
В том случае, если в двух хромосомах разрывы проходят в околоцентромериых районах, два несущих центромеру фрагмента могут объединиться в одну хромосому таким образом, что цептромерные районы этих фрагментов объединяются в одну центромеру, функционирующую как одно целое. Ацентрические фрагменты (по одному плечу от каждой хромосомы) в этом случае рано или поздно теряются из ядра.
Так как в этом случае из ядра теряется по одному плечу от каждой перестроенной хромосомы, то далеко не каждый раз после перестройки возникает жизнеспособная клетка. Однако в некоторых случаях такая делеция может не приводить к летальному исходу и даже не изменять заметно фенотип носителя. Это происходит в том случае, если в перестройке принимают участие акроцентрические хромосомы. Большие плечи и центромерные районы перестроенных хромосом в этом случае объединяются в одну хромосому, а короткие плечи, часто со спутниками, теряются. В большинстве случаев описанные у человека транслокации происходят именно таким образом.
Пусть имеются акроцснтрические хромосомы D и G (рис. 29). Предположим, что разрывы пройдут но области центромеры в одной и другой хромосоме и затем объединятся несущие центромеру фрагменты. В результате этого из двух акроцептрлческих хромосом возникает одна субметацентрическая. Кариотип с наличием такой траислокациониой хромосомы можно условно считать сбалансированным, пренебрегая делецией коротких плеч перестроенных хромосом, содержащих гетерохроматические районы, потому что такая делеция часто не сказывается на фенотипе носителя.
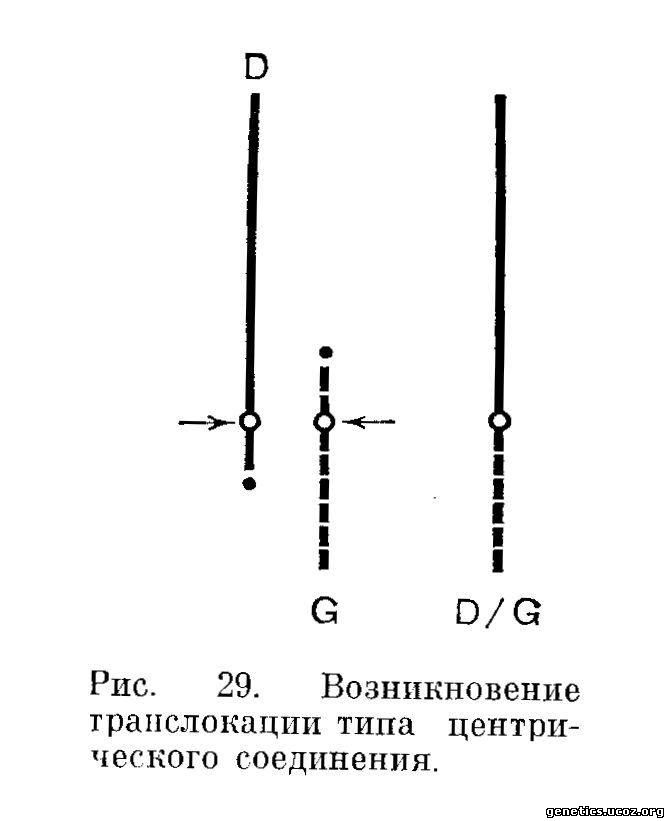
После конъюгации в мейозе возможны три типа расхождения перестроенных хромосом (рис. 30).

В отличие от расхождения из транслокационного креста при сбалансированной трапслокации в данном случае всегда к одному полюсу будет идти одна хромосома, а к другому — две. В том случае, если к одному полюсу веретена деления направится трапслокационная хромосома D/G, а к другому — нормальные гомологи принявших участие в перестройке хромосом, возникнут две гаметьт, одна из которых будет иметь нормальный набор, а другая — условно сбалансированный. В остальных двух теоретически возможных типах расхождения будут возникать по две несбалансированные гаметы. Половина из них будет нести делецию и половина — дупликацию по одной, и притом целой, хромосоме из двух принявших участие в перестройке. Так, если к одному полюсу отойдет хромосома D/G и нормальная хромосома G, то после слияния такой гаметы с гаметой второго (нормального) родителя возникнет кариотип, по существу, трисомичный по хромосомам пары G.
Транслокационный синдром Дауна в большинстве случаев возникает именно таким образом. В этом случае, как и при перицентрической инверсии, реципрокной и нереципрокнои транслокацип, фенотипический носитель соалансированпой перестройки может продуцировать известное количество несбалансированных гамет, что приводит к появлению пораженного потомства.
Однако в отличие от трапслокации между гомологичными хромосомами в перечисленных выше случаях возникают и сбалансированные по перестройке гаметы, дающие у потомства кариотип, аналогичный родительскому. Поэтому такие перестройки могут передаваться в сбалансированном состоянии теоретически через неограниченное число поколений, приводя время от времени к появлению пораженного потомства.
Важнейшим достижением биологии в первой четверти XX века является хромосомная теория наследственности, согласно которой преемственность свойств организмов в ряду поколений определяется преемственностью их хромосом.Хромосомы представляют собой сложноорганизованные нитевидные структуры клеточного ядра, построенные из ДНК и основного белка (гистон) . Хромосомные нити дифференцированы по длине на специфические участки — хромомеры, теломеры, центромеры, гетерохроматические и эухроматические районы, обнаруживаемые в световом микроскопе. Характерным и важнейшим свойством хромосомы является ее способность к спирилизации и деспирализации, определяющая се преобразования в митозе и мейозе и контролирующая ее генетическое функционирование.
Хромосомы выполняют две функции:
1) они точно удваиваются (репродуцируются) перед каждым делением клетки, обеспечивая преемственность наследственного материала в ряду поколений клеток и организмов;
2) участки специфически организованной ДНК хромосом (гены) служат матрицей для синтеза информационной рибонуклеиновой кислоты (m-РНК), осуществляющей синтез специфических белков клетки, определяя тем самым наследственный характер обмена веществ и развития организмов.
Число, размеры и форма хромосом (кариотип) строго постоянны и характерны для каждого вида организмов. В ядре соматических клеток со-держится двойной диплоидный набор хромосом: каждая хромосома представлена парой сходных гомологичных хромосом (отцовского и материнского происхождения).
Постоянство хромосомных наборов (кариотип) в ряду клеточных поколений обеспечивается универсальными закономерностями митоза, в результате которого после удвоения хромосом в делящейся клетке к противоположным полюсам отходят такие же хромосомные наборы, какие были в исходной материнской клетке. Постоянство хромосомных наборов в ряду поколений организмов обеспечивается универсальными закономерностями метоза, центральным событием в котором является конъюгация гомологичных хромосомв профазе мейоза. В анафазе I мейотического деления конъюгировавшие хромосомы разъединяются и расходятся в противоположные клетки (редукция хромосомного набора). В результате мейоза развиваются гаметы, содержащие одиночный гаплоидный набор хромосом. В процессе оплодотворения происходит объединение гаплоидных ядер мужской и женской гаметы (кариогамия) и возникает зигота с диплоидным набором хромосом, дающая начало развитию нового организма.
Повреждения механизмов митоза или мейоза влекут за собой нарушения закономерностей распределения хромосом между сестринскими клетками, в результате чего возникают гаметы или соматические клетки с анормальными численно хромосомными наборами: полиплоидными (при умножении полных хромосомных наборов) или анэуплоидными (при добавлении или утере одной или нескольких хромосом в наборе).
Действие на клетки различных мутагенов (ионизирующая радиация, химические агенты, вирусы) сопровождается разрывами хромосом и часто соединением разорванных участков в новом порядке. Так возникают структурные изменения хромосом (инверсии, транслокации, дупликации и делеции). Численные и структурные изменения хромосом у человека лежат в основе ряда заболеваний.
В настоящее время всеобщее внимание приковано к генетическим событиям, развертывающимся на молекулярном уровне. В основе многих заболеваний и дефектов развития человека лежат механизмы, работающие на микроскопическом уровне, — это нерасхождение хромосом, различные типы хромосомных перестроек, сопровождающиеся нарушением хромосомного баланса в ядре, дифференциальная гетерохроматизация, сопровождающаяся инактивацией генов и компенсирующая нарушения в этом балансе, и др. Таким образом, молекулярная генетика, раскрывая механизмы транскрипции и трансляции наследственной информации, не снимает огромного значения знаний в области закономерностей строения и поведения хромосом на микроскопическом уровне у растений, животных и человека.