Значительное количество экспериментальных исследований по выяснению действия вирусов на хромосомы проведено на культурах различных клеток животных и человека. Большинство работ проведено с вирусом герпеса; далее идут вирус кори, краснухи, инфекционного гепатита, аденовирус 12, вирус Сендай и штаммы вируса саркомы Рауса.
В отличие от противоречивых результатов некоторых исследований in vivo, во всех работах, проведенных на культурах клеток, инфицированных вирусом, было установлено повреждающее действие данного агента на хромосомы. Большое значение имеет обнаружение Nichols с соавторами различий в этом отношении двух линий вируса саркомы Рауса: при инфицировании культуры лейкоцитов человека Sr-линией этого вируса наблюдались многочисленные клетки с разрывами хромосом, тогда как линия Бриана того же вируса никакого повреждающего действия на хромосомы не оказывала. Эти данные проливают некоторый свет на противоречивые результаты, полученные в разные годы при исследовании действия одного и того же вируса (Makino и др.,1966).
Многочисленными исследованиями установлено, что под действием некоторых вирусов культуры клеток претерпевают морфологическую и физиологическую трансформацию, связанную с изменениями в хромосомных наборах.
Вакуолизирующий вирус обезьяны SV-40 вызывает in vitro трансформацию клеток китайского и сирийского хомячка и клеток человека. В трансформированных SV-40 культурах клеток сирийского хомячка наблюдаютея многочисленные хромосомные перестройки и полиплоидные клетки (Black, Rowe, 1963). Клетки китайского хомячка, которые претерпевали трансформацию под влиянием SV-40 и приобрели злокачественные свойства, сохранили нормальный кариотип (Yerganian, 1964).
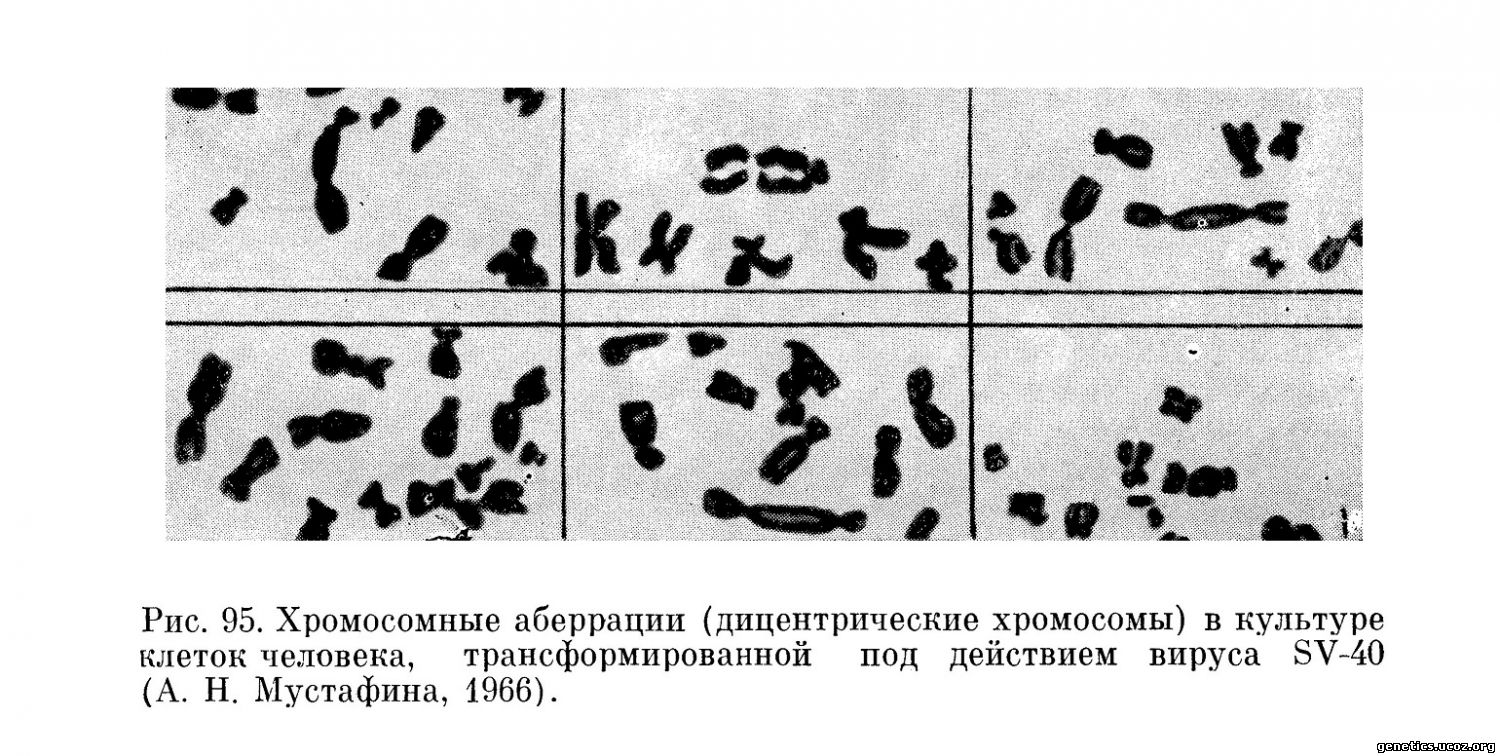
Многочисленные и разнообразные изменения под воздействием вируса SV-40 претерпевают хромосомы в процессе трансформации клеток человека (Koprowski и др., 1962; Morrhead и др., 1963; М. П. Чумаков и др., 1964; А. Н. Мустафина, 1966).
Как известно, этот латентный онкогенный вирус почек обезьян может контаминировать некоторые вирусные вакцины, применяемые на людях, и обнаружение трансформирующего его действия на клетки человека имело большое значение.
Среди обширных исследований, развернувшихся в этой области, специальные наблюдения были проведены для выяснения действия вируса SV-40 на хромосомный аппарат клеток (Moorhead, Saksela, 1963, 1965; Yerganian и др., 1962; Torado и др., 1963; Wolman и др., 1964; А. Н. Мустафина, 1966) (рис. 95).
Вирус саркомы Рауса вызывает злокачественные опухоли не только у цыплят и кур, но также у многих млекопитающих, включая крысу, мышь, морскую свинку, хомячка и кролика. Nichols с сотрудниками (1965) провели серию исследований хромосомных наборов в процессе трансформации диплоидных клеток крысы под влияним вируса саркомы Рауса in vivo и in vitro.
При исследовании клеток опухоли in vivo в нулевом и ранних пассажах ни количественных, ни качественных изменений в хромосомных наборах обнаружено не было. Однако в 20-м пассаже в результате разрывов и перестройки хромосом появились новые хромосомные типы. В 50-м пассаже наблюдалось большое количество перестроенных хромосом и полиплоидных клеток. Сходным образом вели себя клетки первичной опухоли в культуре: в процессе трансформации отмечалось большое количество различных типов оберраций хромосом. Затем устанавливалась более или менее стабильная клеточная линия с диплоидным числом хромосом, но со структурными изменениями. Эти клетки при их инъекции новорожденным крысам вызывали опухоли у 100% животных. Инфицирование культуры клеток крысы вирусом саркомы Рауса также значительно повышало число клеток с разрывами хромосом.
Кроме повреждений хромосом в клетках, инфицированных вирусом, обнаруживаются и другие нарушения, касающиеся изменений характера митотической спирализации хромосом, анормальностей митоза и изменения клеточного цикла.
Резкое сокращение хромосом, сходное с действием, вызванным колхицином, описано при инфицировании клеток вирусом простого герпеса, опоясывающего лишая и желтой лихорадки (Воігоп и др., 1966; Tanzer и др., 1964; Benyesh-Melnick, 1964).
Ненормальная деспирализация хромосом, приводящая метафазные хромосомы в состояние нитей профазы, обнаружена при действии вирусов простого герпеса и опоясывающего лишая на диплоидные линии клеток китайского хомячка и эмбрионов легкого человека (Stich, Hsu, 1963; Stich и др., 1964а), а также при действии вируса Сендай на хромосомы в диплоидных клетках человека и культуры Hela (Aula, Saksela, 1966).
Анормальные митозы — отставания хромосом, многополюсные митозы, многоядерные и полиплоидные клетки — установлены как результат действия целого ряда вирусов: простого герпеса (Воігоп и др., 1966; Tanzer и др., 1964), опоясывающего лишая (Benyesh-Melnick, 1964), вирусов паротита и клещевого энцефалита (С. Я. Залкинд, 1966), вируса SV-40 (Koprowski и др., 1962; М. П. Чумаков и др., 1964), кори (Fjeed, Holtermann, 1962), аденовирусов 12 и 18 (Stich и др., 1963, 1964),
Имеются многочисленные указания на заметное повышение при вирусных инфекциях числа анэуплоидных клеток (Moorhead, Saksela, 1965; А. Н. Мустафина, 1966, Ю. Я. Керкис и др., 1967; Gripenberg, 1965).
Все эти нарушения с несомненностью указывают на повреждение ахроматического аппарата митоза в клетке, инфицированной вирусом.
В культуре лимфоцитов периферической крови больных корью наблюдались также слияние клеток и их ядер (Nichols и др., 1965), фрагментация ядер и нарушения в числе хромосом, которые могут быть объяснены их частичной эндоредупликацией (Gripenberg, 1965).
Обширные данные свидетельствуют, что многие вирусы оказывают влияние на клеточный цикл.
Стимуляция митозов отмечена в клетках, инфицированных онкогенными вирусами SV-40 (Koprowski и др., 1963) и вирусом саркомы Рауса (Rubin, Temin, 1958). Имеются указания на то, что вирусы герпеса, полиомиелита и осповакцины также повышают митотическую активность клеток (Grey и др., 1958; С. С. Николау, 1955; С. Я. Залкинд и Л. Г. Степанова, 1960). Высказано предположение, что повышение митотической активности в этих случаях связано с защитной стимуляцией обмена веществ.
Значительно чаще действие вируса сопровождается подавлением митотической активности клеток, вплоть до полного прекращения клеточных делений. Обстоятельными исследованиями установлено, что вирус герпеса блокирует клетку в периодах S и G2, по-видимому, подавляя синтез ДНК и разрушая ахроматический аппарат клетки (Stoker и др., 1959, Wildy и др., 1961).
Ингибирование митозов наблюдается также при инфицировании клеток вирусами паротита (Walter, 1960), ветряной оспы (Aula, 1963), вирусом Сендай (Saksela и др., 1965), саркомы Рауса (Nichols и др., 1964), а также вирусом кори (Nichols и др., 19646).
Относительно природы ингибирования митозов в результате действия вируса имеется ряд предположений: рассматривается возможность повреждения вирусом ядрышка, имеющего большое значение в пусковом механизме митоза (Wheelock и др.,, 1961), указывается на возможность подавления вирусом синтеза макромолекул, необходимых для клеточного деления (Tobey и др., 1965), подавления синтеза белка, РНК и ДНК.
Таким образом, действие многих вирусов сопровождается нарушением клеточного метаболизма, приводящим в некоторых случаях к стимуляции митоза. Однако при инфицировании клеток действие большинства вирусов сопровождается подавлением митотической активности клеток.