Кариотип. Описанная выше методика исследования хромосом человека, обеспечила не только установление их числа, в отношении которого данные многочисленных цитологов оказались тождественными, но с помощью этой методики были выявлены также и структурные особенности аутосом и половых хромосом человека, определяемые их размерами, положением центромерного участка, наличием вторичных перетяжек и спутников.
Уже Chi и Giles (1959) полагали, что в хромосомном наборе человека можно индивидуально идентифицировать каждую пару гомологичных хромосом. Данные этих авторов показали также, что гомологичные аутосомы в клетках различных тканей того же самого или различных индивидуумов не отличаются существенно между собой ни относительными длинами, ни положением центромерного участка. Единственное различие между хромосомными наборами двух полов у человека состоит в половых хромосомах: в диплоидном наборе женщины имеются две одинаковые хромосомы X, в диплоидном наборе мужчины содержится одна, такая же как у женщины, хромосома X, и маленькая Y-хромосома. Полученные разными цитологами независимо друг от друга данные по установлению структуры хромосом человека так же хорошо совпадали, как и данные о числе хромосом.
Таким образом, кариотип человека как совокупность данных о числе, размерах и структуре хромосом человека установлен в настоящее время с большой точностью. Характерные особенности строения каждой хромосомы определяются, как известно, положением в ней центромеры важнейшего ее органоида, который в митозе соединяется с нитями веретена и определяет расхождение сестринских хроматид к противоположным полюсам клетки. В районе центромеры хромосома сужена, две ее хроматиды сближены, и этот район образует в теле хромосомы первичную перетяжку. По положению центромеры и первичной перетяжки среди хромосом человека различают метацеытрические хромосомы, у которых центромера расположена в середине хромосомы (медианно) и делит ее на два равных по длине плеча (рис. 53, а), субметацентрические хромосомы, в которых центромера расположена суомедианно и делит хромосому на два плеча неравной длины (рис. 33, б), и акроцентрические, у которых центромера расположена почти на конце хромосомы (терминально), отделяя от длинного плеча очень короткое; у некоторых хромосом на коротком плече на красящейся тонкой нити располагается маленькое хроматическое тельце — спутник (рис. 33, в).

Плечи некоторых хромосом содержат перетяжки, называющиеся «вторичными». 46 хромосом диплоидного набора человека составлены 23 парами гомологичных (отцовского и материнского происхождения) хромосом: 22 пары аутосом + половые хромосомы XX (у женщин) или XY (у мужчин). Гомологичные хромосомы, как правило, сходны между собой в размерах и строении, хотя могут встречаться некоторые отклонения от каждого из этих показателей.
Интенсивное исследование хромосом человека в лабораториях разных стран сопровождалось созданием цитологами нескольких различных систем классификации и номенклатуры хромосом человека. Возникшие при этом затруднения при идентификации хромосом, обозначаемых исследователями разными символами, привели к настоятельной необходимости унификации номенклатуры хромосом человека. Важнейшая работа по созданию общей системы обозначения хромосом человека была проведена в 1960 г. специальной комиссией, собравшейся в Денвере, в Университете штата Колорадо (США), при поддержке американского канцерологического общества. В состав комиссии вошло 14 цитологов, опубликовавших в 1960 г. данные о кариотипе человека (Book, Chu, Ford, Fraccaro, Harnden, Hsu, Hungerford, Jacobs, Lejeune, Levan, Makino, Puck, Robinson, Tjio). Кроме цитологов, в комиссию вошли 3 арбитра-генетика. Результаты работы этой комиссии опубликованы в ряде журналов в виде документа, обозначенного как «Стандартная система номенклатуры митотических хромосом человека» (Robinson, 1960).
Согласно этому документу, термин «кариотип» рекомендуется применять к систематизированному набору хромосом единичной клетки, выполненному с помощью рисунка или путем вырезывания хромосом из микрофотографии (рис. 34, 35). При этом имеется в виду, что хромосомы данной клетки типичны для индивидуума.
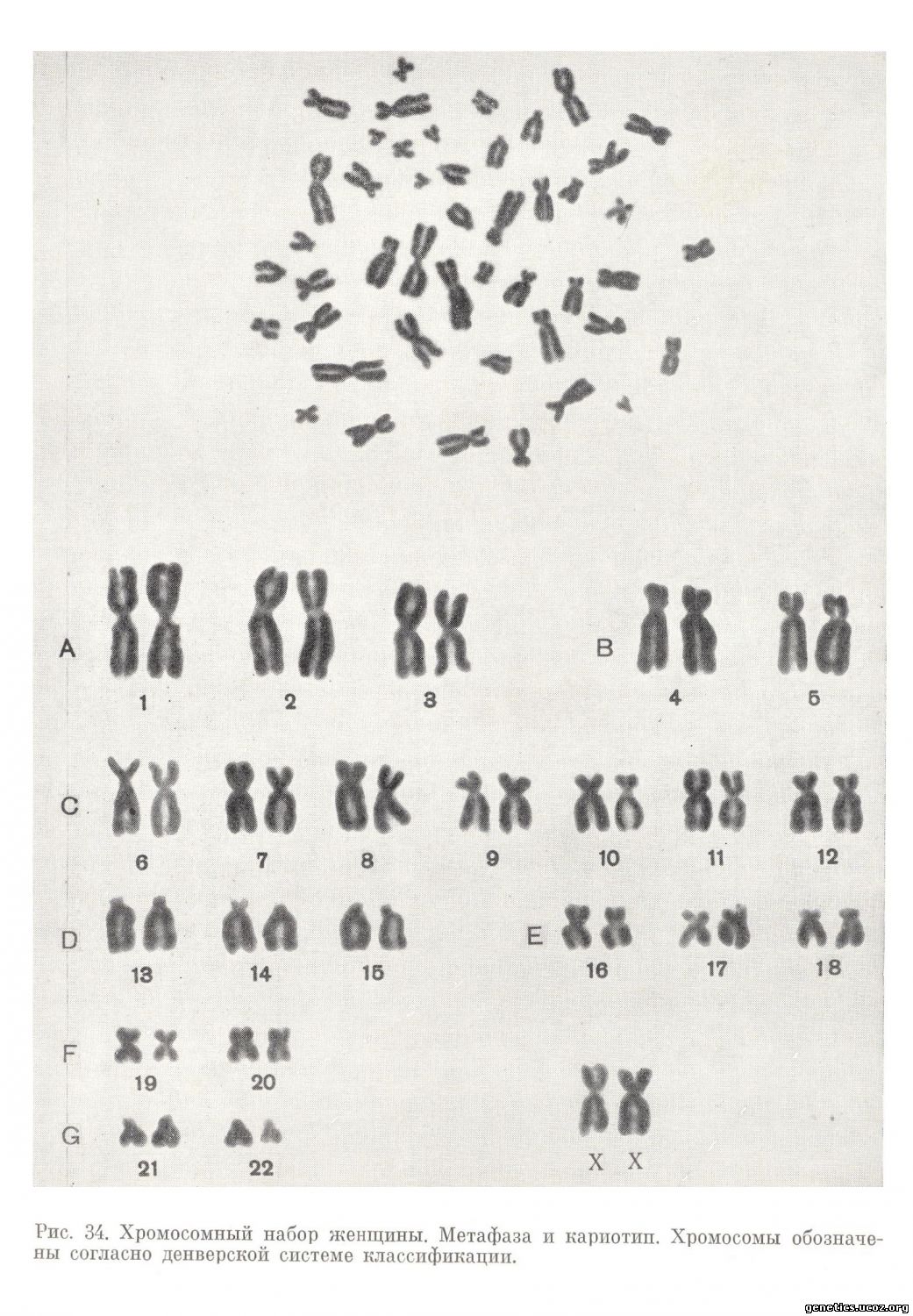

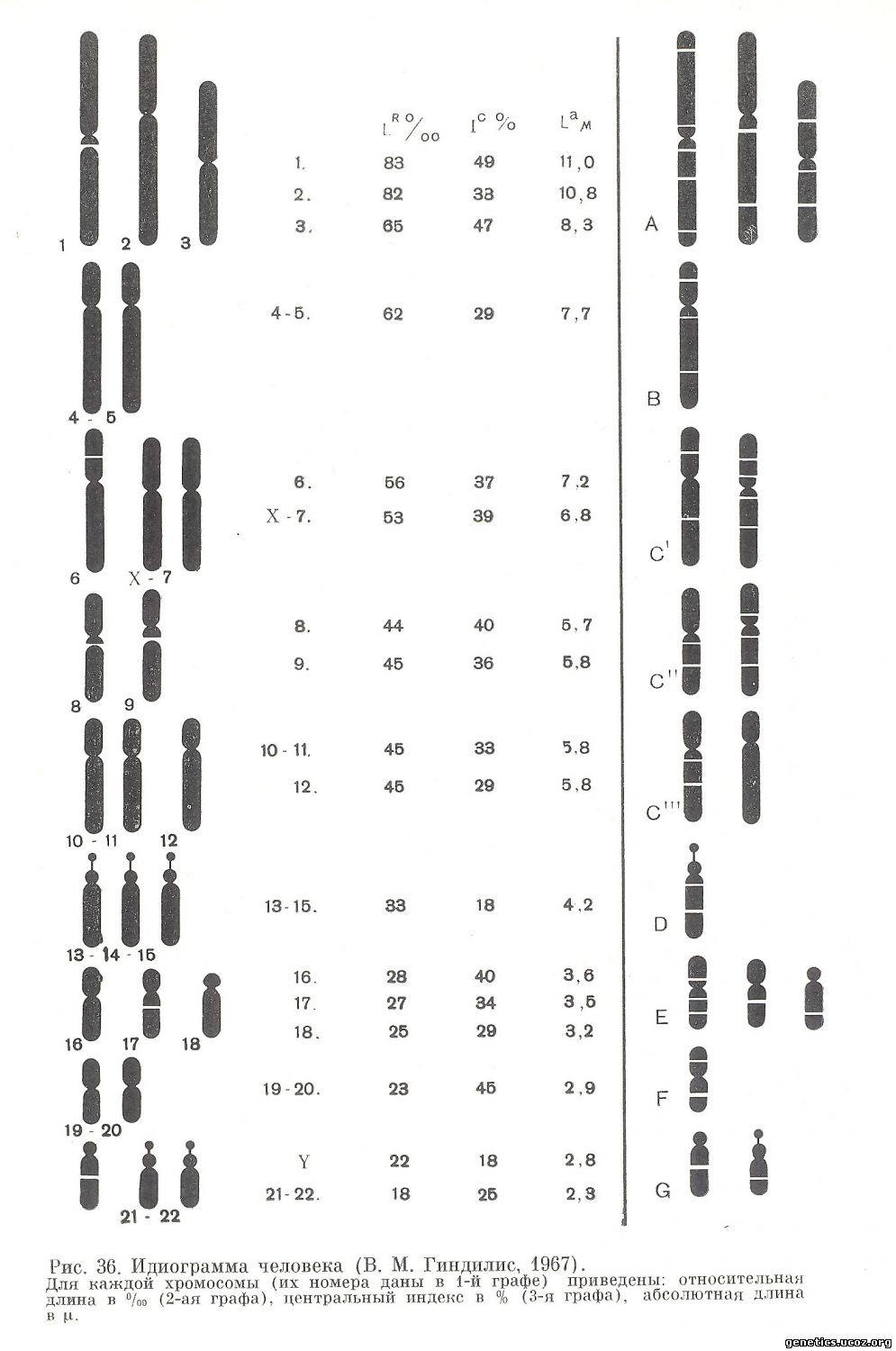
Термин «идиограмма» сохраняется для представления кариотипа в виде схемы, построенной на основании измерений хромосом нескольких клеток (рис. 36) . Хромосомы пронумерованы серийно от 1 до 22 в соответствии с их длиной, а также с другими особенностями их строения, допускающими идентификацию. Половые хромосомы не имеют номеров и обозначаются как X и Y.
В 1963 г. в Лондоне состоялась вторая конференция по стандартизации данных о нормальном кариотипе человека. На этой конференции было констатировано, что номенклатура хромосом человека, разработанная на Денверской конференции, выдержала испытание временем. Вместе с тем на Лондонской конференции было обращено внимание на важность для идентификации и индивидуализации хромосом таких важных их характеристик, как наличие вторичных перетяжек и особенности рисунка инкорпорации тимидина-Н3, определяемого методом авторадиографии. Лондонская конференция (1963) внесла некоторые уточнения в номенклатуру хромосом, а Чикагская конференция (1966) разработала систему обозначений нормальных и измененных наборов хромосом человека. К настоящему времени информация о структуре хромосом человека расширена данными о положении в них вторичных перетяжек (Saksela, Moorhead, 1962; В. М. Гиндилис, 1966) (рис. 37, а, б); 22 аутосомы распределены в семь групп, различия между которыми легко определимы.
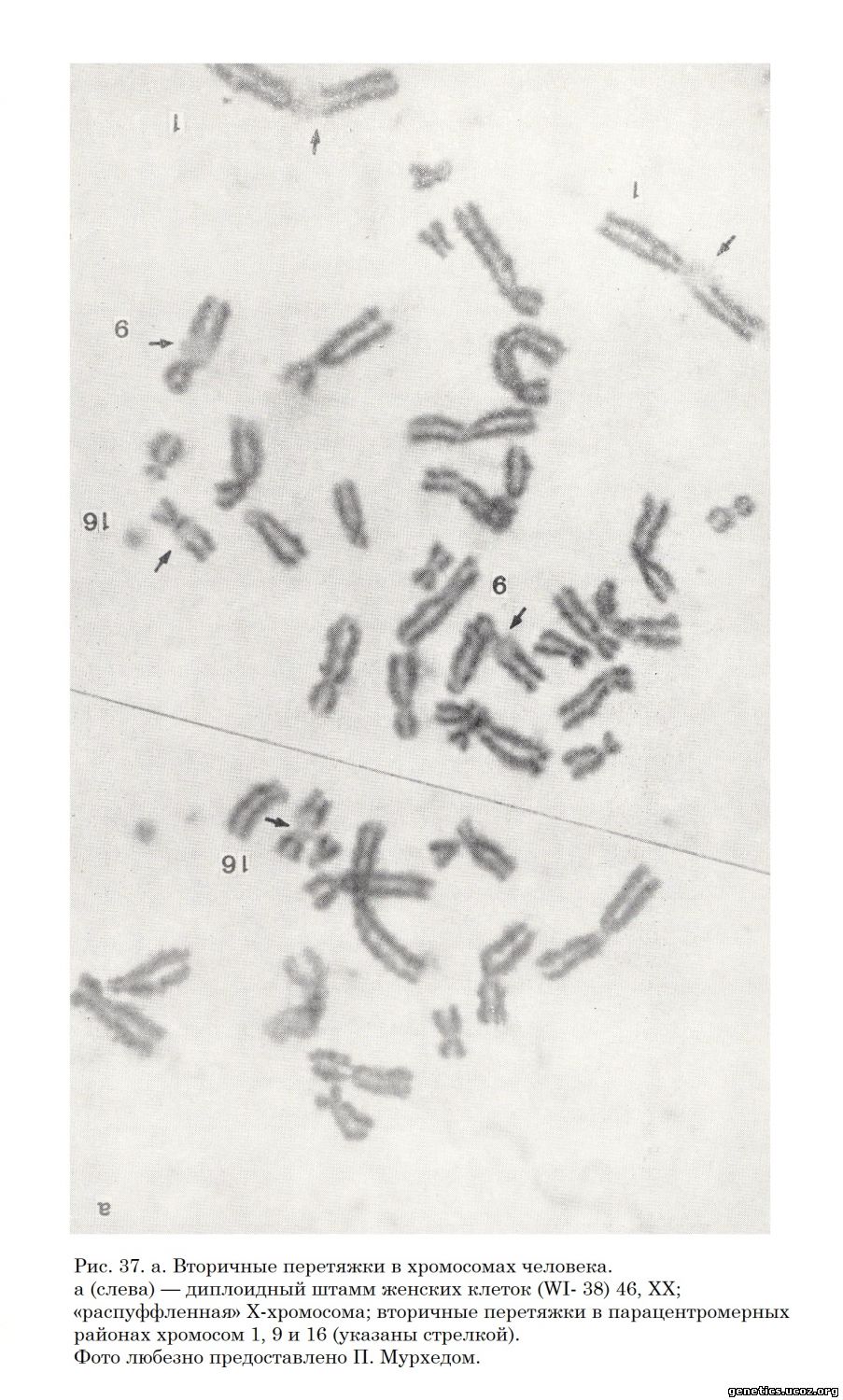
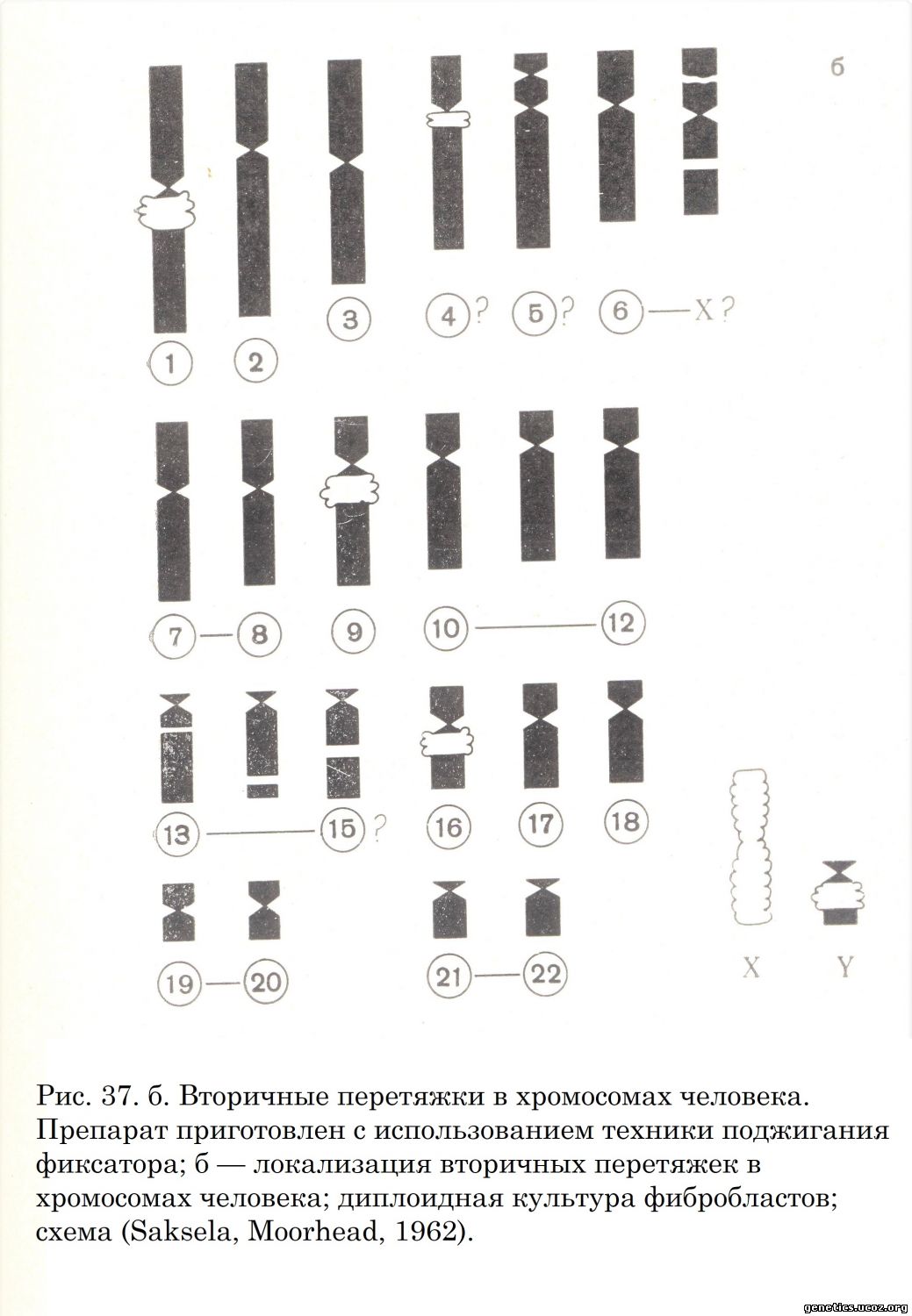
Ниже приводится классификация и номенклатура хромосом человека, составленная на основе объединенных данных Денверской и Лондонской конференций и дополненная данными о положении вторичных перетяжек.
Группа 1—3. Крупные хромосомы с почти медианными центромерами в хромосомах 1 и 3 и субмедианной в хромосоме 2. Эти три хромосомы хорошо отличаются одна от другой размерами и положением центромеры. Хромосома 1 содержит вторичную перетяжку в субцентромерном районе одного из плеч (более длинном).
Группа 4—5. Крупные хромосомы с субмедианными центромерами. Хромосомы этих пар ни морфологически, ни размерами не различаются между собой. В редких случаях в хромосомах этой группы обнаруживается вторичная перетяжка в длинном плече вблизи центромеры
Группа 6—12. Хромосомы средних размеров с субмедианными центромерами. Х-хромосома сходна с самыми длинными хромосомами этой группы — с 6-й и 7-й, от которых ее трудно отличить. Четыре из хромосом как 6, 7, 8 и 11. Х-хромосома относится к этой группе. Три хромосомы более субметацентричны, их следует нумеровать как 9, 10 и 12. Эта большая группа хромосом представляет наибольшие трудности при идентификации индивидуальных хромосом. Хромосома 6 имеет вторичную перетяжку в середине короткого плеча. Хромосомы 8 и 9 содержат вторичную перетяжку в длинном плече вблизи центромеры.
Группа 13—15. Хромосомы средних размеров с почти терминальными центромерами («акроцентрические» хромосомы). Все три пары хромосом являются потенциально спутничными, но не в каждой клетке и не у всех индивидуумов спутники обнаруживаются на всех хромосомах. Чаще всего спутники обнаруживаются на одной или двух парах. Между собой три пары хромосом морфологически неразличимы. Длинное плечо одной из этих хромосом имеет вторичную перетяжку вблизи центромеры.
Группа 16—18. Довольно короткие хромосомы с почти медианными (хромосома 16), субмедианными (хромосома 17) и субтерминальными (хромосома 18) центромерами. Хромосомы 16 и 17 содержат в длинном плече вблизи центромеры вторичную перетяжку.
Группа 19—20. Короткие хромосомы с почти медианными центромерами. Между собой неразличимы.
Группа 21—22. Очень короткие акроцентрические хромосомы. Обе пары хромосом потенциально имеют спутники. Однако обычно лишь две хромосомы имеют на коротком плече хорошо выраженные спутники. На других хромосомах спутник выявляется реже. Между собой эти две хромосомы неразличимы.
Y-хромосома. Маленькая акроцентрическая хромосома; сходна по размерам и форме с хромосомами 21 и 22, но не имеет спутников. Отличается от хромосом 21 и 22 также большей пикнотичностью и сближенными хроматидами длинного плеча. В середине длинного плеча имеет вторичную перетяжку.
Метод построения идиограмм и кариотипа, предложенный Денверской комиссией, был подвергнут критике Patau (1960), который указал на субъективность подбора пар гомологичных хромосом на основе сходства их по форме и длине. Степень спирализации гомологичных хромосом может быть различной как в разных клетках, так и в той же самой клетке, поэтому подбор гомологичных хромосом по длине может привести к неправильной идентификации пар.
Эти соображения Patau весьма обоснованы, так как специальные исследования у ряда объектов выявили закономерную гетероцикличность родительских хромосом в системе диплоидного ядра (А. А. Прокофьева-Бельговская, 1948) и указания на этот феномен имеются в ряде более поздних работ (Ołino, Hauschka, 1960; Гиндилис, 1966; А. А. Прокофьева-Бельговская и С. И. Слезингер, 1965). Patau предложил классифицировать большинство хромосом человека лишь по группам, в соответствии с их размерами и положением центромеры: группа А — крупные метацентрические (1, 2 и 3), группа В — крупные субметацентриче- ские (4 и 5), группа С — средние субметацентрические (6—X—12), группа D — крупные акроцентрические (13, 14, 15), группа Е — маленькие субметацентрические (16, 17, 18), группа F — маленькие метацентрические (19 и 20), группа G — маленькие акроцентрические (21, 22). Кроме группы А, в других группах хромосомы либо совсем не идентифицируются, либо идентифицируются с трудом.
Некоторые исследователи в настоящее время используют двойную номенклатуру, обозначая хромосомы, согласно классификации Денверской комиссии, с буквенными уточнениями, предложенными Patau.