Репродукция аутосом в культуре лейкоцитов периферической крови изучена наиболее полно. Хотя в отношении единичных хромосом результаты исследований отдельных авторов противоречивы, в отношении большинства хромосом данные различных исследователей находятся между собой в хорошем соответствии (Gilbert и др., 1902; Schmid, 19G3; German, 1964, 1965; Kikuchi, Sandberg, 1964; N. Bianchi, M. Bianchi, 1965).
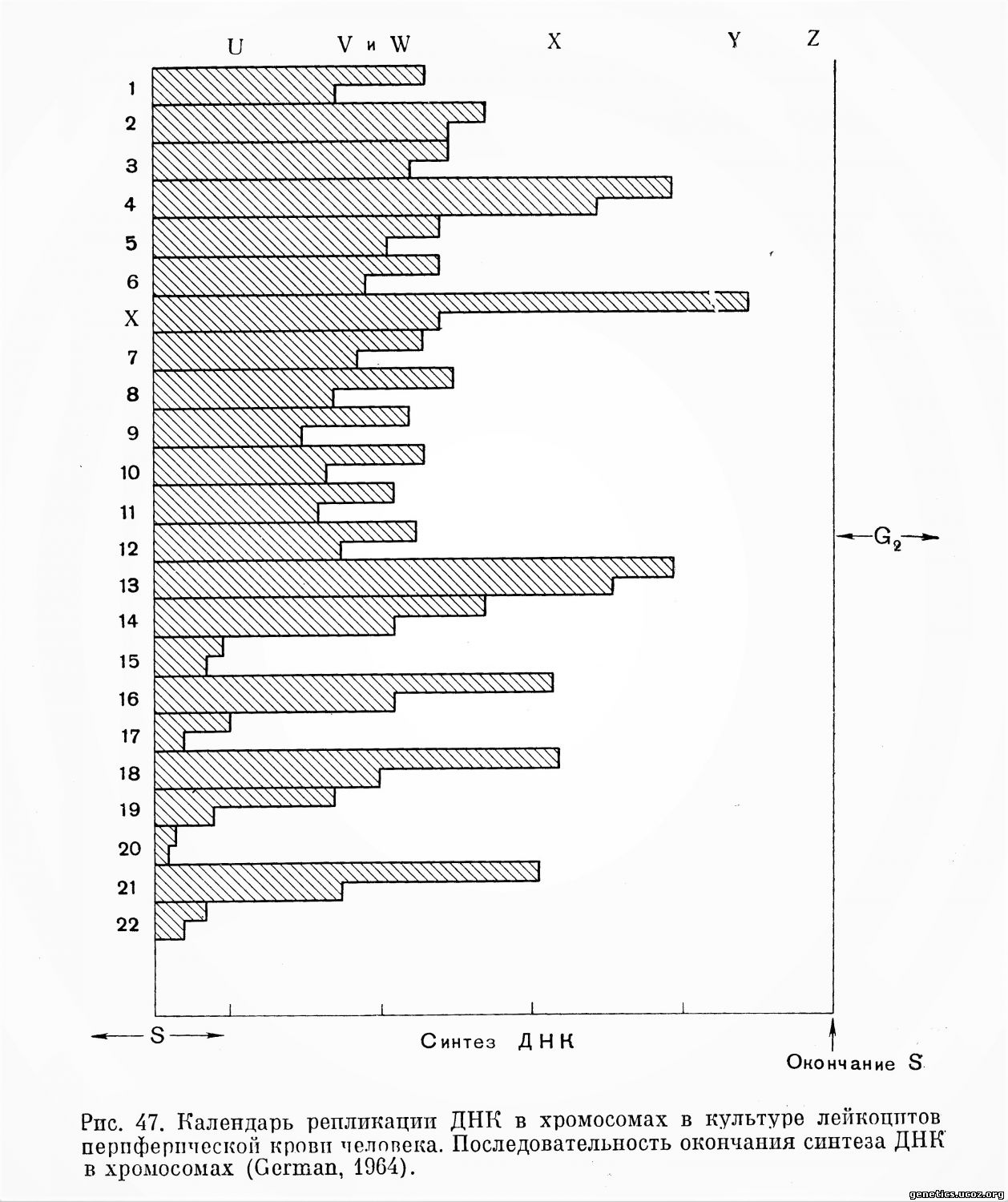
Календари последовательности репродукции хромосом человека в культуре лейкоцитов по материалам разных авторов представлены на рис. 47, 48.

При использовании авторадиографии для идентификации хромосом предполагается, что индивидуальные хромосомы обладают специфическими особенностями репродукции по длине и времени окончания этого процесса.
Предполагается также, что гомологичные хромосомы сходны между собой в особенностях репликации (рисунок и интенсивность мечепия в конце периода S). В соответствии с этим наиболее обстоятельные исследования (Gilbert и др., 1962; Schmid, 1963; Muldal и др., 1963; German, 1964, 19G5; German и др., I960) устанавливают ряд особенностей репродукции отдельных хромосом и групп хромосом в конце периода синтеза.
Хромосома 1. Теломерные районы и значительная часть обоих плеч представляют собой участки, заканчивающие репродукцию рано. Позднее реплицируется проксимальный район одного из плеч (более длинного) и центромерный район. В самом конце периода S «горячим» участком в хромосоме 1, является, по-видимому, район вторичной перетяжки.
Хромосома 2. Заканчивает репликацию заметно позднее хромосомы 1. Теломерные районы заканчивают репликацию ранее других: позднее реплицируются участки, расположенные в обоих плечах у центромеры. В самом конце периода S эта хромосома уже не имеет «горячих» участков.
Хромосома 3. Относится к сравнительно поздним репликантам. Ранее всего заканчивают репродукцию теломерные районы. Наиболее поздно реплицируется центромерный район.
Хромосомы 4—5. Хромосомы этой группы на основании морфологических признаков и количественных показателей между собой неразличимы. Обе пары хромосом относятся к поздним репликантам и различаются по времени окончания синтеза ДНК: в длинных плечах двух хромосом этот процесс уже заканчивается, в то время как в двух других он интенсивно продолжается по всей длине хромосомы. Условно поздний репликант обозначен как хромосома 4 и ранний — как хромосома 5. Короткие плечи хромосом 4—5 заканчивают репликацию поздно.
Хромосомы 6—12. Хромосомы этой группы не имеют специфических,, выделяющихся особенностей мечения. В конце периода S в хромосоме 6 интенсивное включение тимидина-Н3 обнаружено в центромерном районе.
Хромосомы 13—15. Три пары хромосом неразличимы между собой морфологически и по количественным показателям. Они различаются особенностями репродукции. Две хромосомы этой группы, наиболее короткие и потому обозначаемые как хромосомы 15 (D3), относятся к группе наиболее ранних репликантов хромосомного набора. Две хромосомы относятся к группе наиболее поздних репликантов, они обозначены как хромосомы 13 (D1). Хромосома 14 (D2) в отношении времени окончания репродукции занимает промежуточное положение. Имеются указания, что у двух хромосом — поздних репликантов (D1) в конце периода S «горячим» участком является дистальная половина длинного плеча, тогда как у хромосомы D2 таковым является короткое плечо и центромерный район (Yunis, 1964; German и др., 1966). Согласно другим данным, наиболее часто самый поздний реплицирующийся участок — середина длинного плеча этих хромосом (German, 1964).
Хромосома 16. Данные в отношении особенностей репродукции этой хромосомы противоречивы. Как правило, она не регистрируется среди поздних репликантов.
Хромосома 17. Относится к хромосомам, наиболее рано заканчивающим репродукцию.
Хромосома 18. Поздно репродуцирующаяся хромосома.
Хромосомы 19—20. Обе пары хромосом заканчивают репродукцию очень рано и неразличимы между собой по рисунку включения тимидина-Н3.
Хромосомы 21—22. Две пары хромосом этой группы не различаются между собой по морфологическим и количественным показателям. Обе пары относятся к ранним репликантам, Однако их различают по времени окончания процесса репродукции.
В двух хромосомах этот процесс заканчивается заметно раньше, чем в двух других. Какую из них считать 21-й и какую 22-й, единого мнения нет. Данные по трисомии в случаях монголизма показывают, что эти нарушения в группе G связаны с «горячей», наиболее поздно реплицирующейся хромосомой.
Таким образом, в культуре лейкоцитов периферической крови ранними репликантами являются аутосомы 1, 15, 17, 19—20, 21—22, поздно заканчивается синтез ДНК в хромосомах 4—5 (наиболее поздно в хромосоме 4)у 13, 18.
Анализ специфических особенностей репродукции индивидуальных хромосом, как полагают некоторые исследователи, открыл возможность идентифицировать хромосомы в группах В, D и G.
В группе В обнаруживаются две более ранние хромосомы и две — более поздние репликанты (German, 1964а, Ь); они в настоящее время рассматриваются как хромосомы 5 и 4 соответственно.
В группе D установлены две пары хромосом как поздние репликанты (13 и 14) и одна пара — ранние репликанты (15) (Schmid, 1963; German, 1964а, Ь).
В группе G две хромосомы представляют собой более ранние (21?) и две — более поздние (22?) репликанты.
В какой степени подобные характеристики индивидуальных пар хромосом соответствуют их истинной гомологии, в настоящее время сказать трудно. Обширный материал показывает, что гомологичные хромосомы в конце периода синтеза могут отчетливо различаться между собой, будучи одна поздним, а другая — более ранним репликантом.
Однако ряд исследователей уже теперь использует особенности репликации индивидуальных хромосом в группах В, D и G при идентификации их в случаях транслокаций и делеций, а также трисомии сопровождающихся соответствующими заболеваниями.
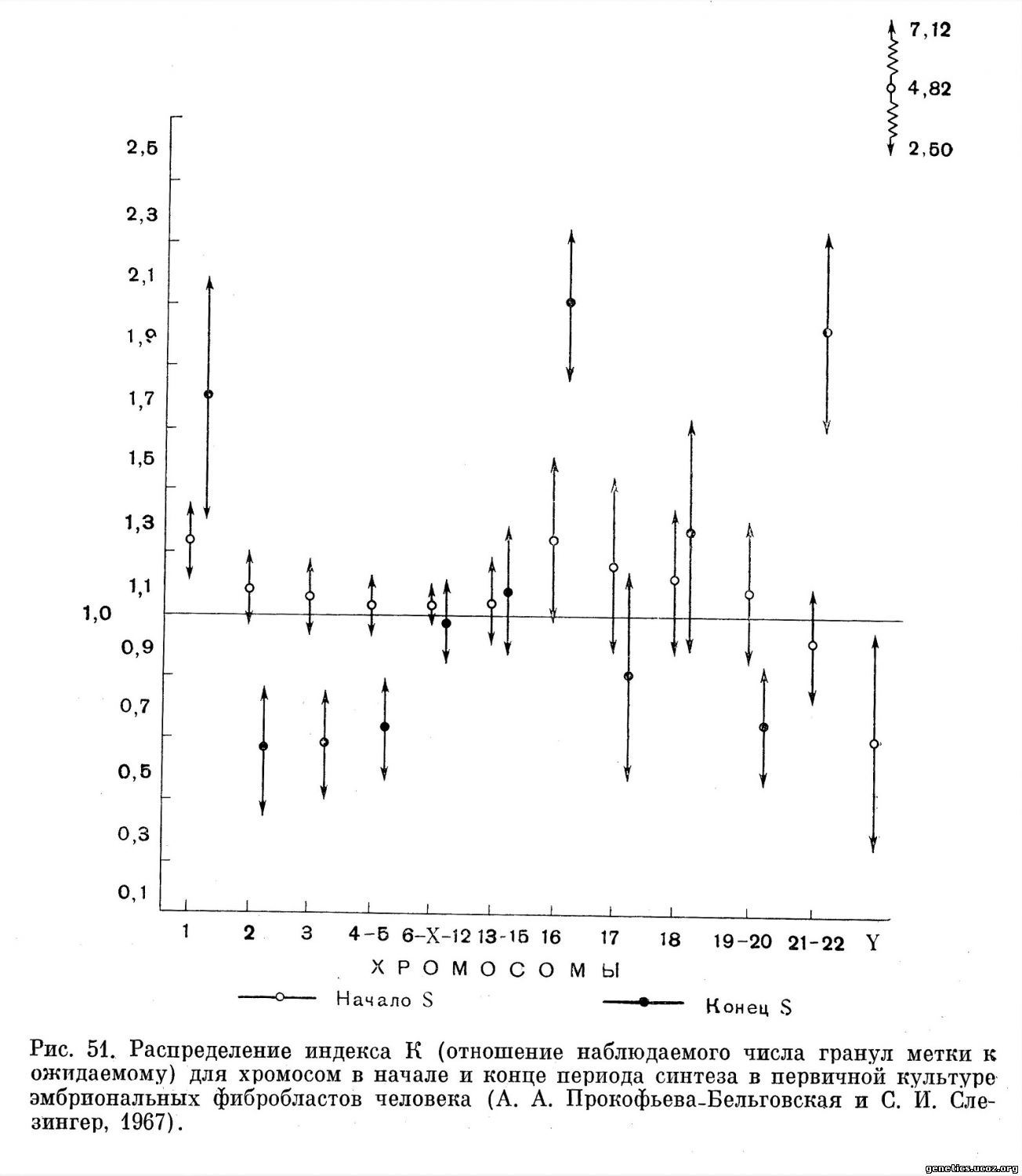
Репродукция хромосом в культуре эмбриональных фибробластов и фибробластов кожи исследована также достаточно обстоятельно. В первичной культуре эмбриональных фибробластов (А. А. Прокофьева-Бельговская и С. И. Слезингер, 1965, 1967; С. И. Слезингер и А. А. Прокофьева-Бельговская, 1965, 1966, 1969) почти все хромосомы вступают в период синтеза более или менее одновременно, но заканчивают репликацию резко асинхронно (рис. 49, 50, 51). В исследовании расчет концентрации метки производили, исходя из относительной длины хромосомы, поэтому можно заключить, что в первичной культуре эмбриональных фибробластов хромосомы различаются между собой в отношении скорости синтеза ДНК и репродукции участков.
Низкая концентрация метки в начале периода S на Y-хромосоме связана, очевидно, с ее поздним вступлением в репликацию. В конце периода синтеза репликация ДНК рано заканчивается в хромосомах 2, 3 и в хромосомах групп 4—5 и 19—20. Поздними репликантами в первичной культуре эмбриональных фибробластов являются хромосомы 1, 16, Y и хромосомы группы 21—22.
В большом соответствии с картиной репродукции хромосом в конце периода синтеза в первичной культуре эмбриональных фибробластов находятся данные о репродукции хромосом в этот же период в сходных типах делящихся клеток — 18—35-й субкультуре эмбриональных фибробластоподобпых клеток (Moorhead, Defendi, 1963) и во 2-й и З-й субкультурах клеток кожи (Atkins, Santesson, 1964, 1966; Atkins, Gustavson, 1964).
Таким образом, асинхронность репликации ДНК хромосом в обоих рассмотренных типах клеток наиболее отчетливо выражена в конце периода синтеза (рис. 51).
Высокая асинхронность в репродукции хромосом человека обусловлена, как показали исследования, различиями участков хромосом в отношении прежде всего срока окончания этого процесса и скорости его протекания.
Длительность процесса репликации ДНК в отдельных хромосомах не зависит от размеров хромосомы; этот факт убедительно доказан исследователями, которые при обработке результатов подсчета гранул метки над хромосомами провели расчет на единицу длины хромосомы (Gilbert и др., 1962; German, 1964; С. И. Слезингер и А. А. Прокофьева-Бельговская, 1965, 1966). Этот анализ показал, что в некоторых длинных хромосомах, например хромосомах 1 и 2, репликация ДНК заканчивается раньше, чем в коротких хромосомах 13, 16, 21. В первичной культуре эмбриональных фибробластов практически одинаковые по длине хромосомы 1 и 2 (=10 ц) заканчивают репликацию ДНК в резко различное время: хромосома 2, вступая в период синтеза несколько позднее хромосомы 1, заканчивает репликацию ДНК значительно раньше, т. е. реплицируется быстрее (С. И. Слезингер и А. А. Прокофьева-Бельговская, 1965, 1966, 1968; А. А. Прокофьева-Бельговская и С. И. Слезингер, 1967).
Большой интерес представляет сопоставление особенностей репродукции одних и тех же хромосом в культурах двух различных типов клеток: культуре лейкоцитов и культуре эмбриональных фибробластов (рис. 52). Сравниваемые культуры отчетливо различаются поведением в конце периода синтеза двух самых крупных хромосом 1 и 2.
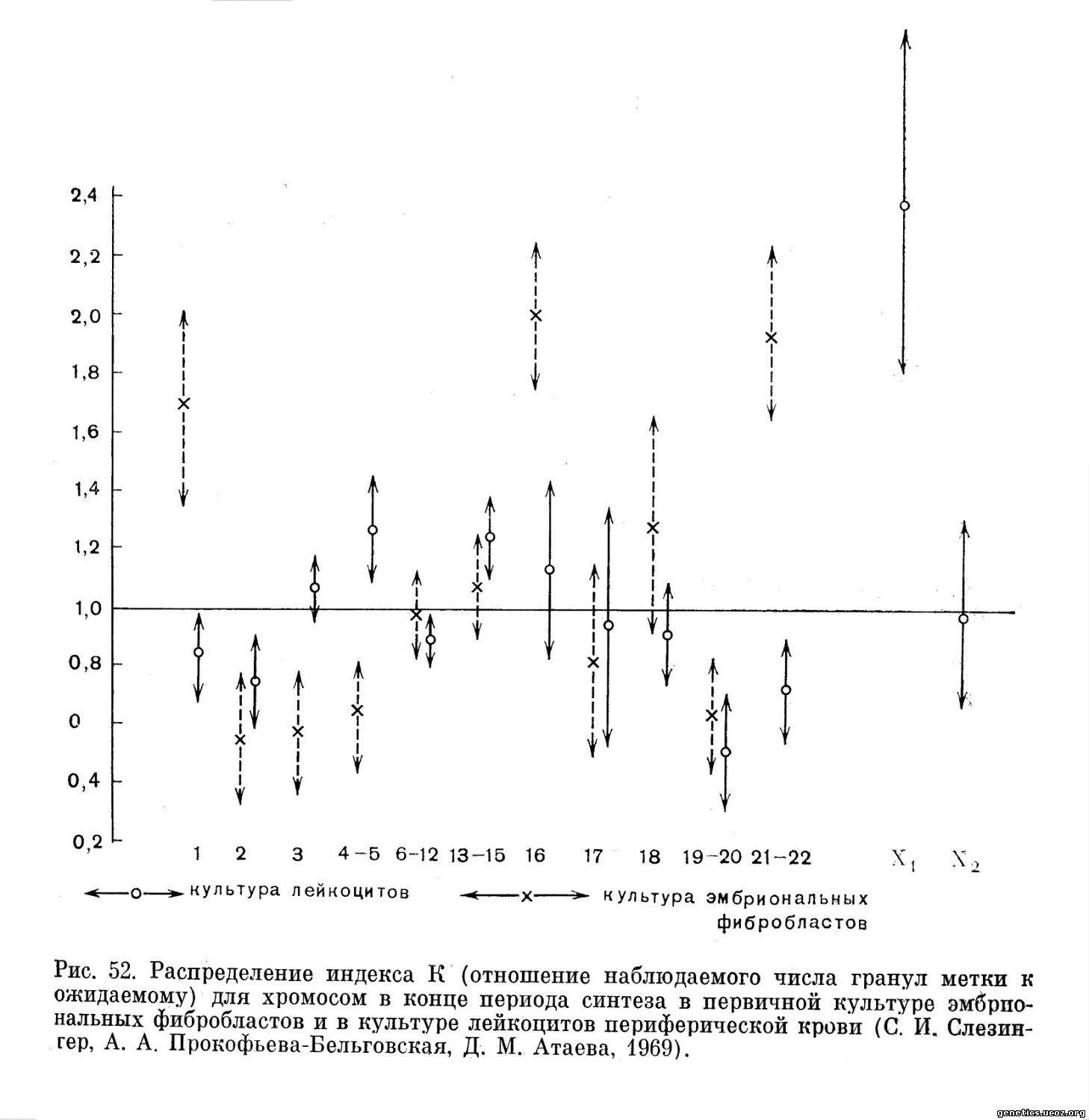
В культуре лейкоцитов рано заканчивает репликацию ДНК хромосома 1 и позднее — хромосома 2 (Gilbert и др., 1962; German, 1964; Kikuchi, Sandberg, 1964; N. Bianchi, M. Bianchi, 1965; С. И. Слезингер и А. А. Прокофьева-Бельговская, 1966; А. А. Прокофьева-Бельговская и С. И. Слезингер, 1967).
В трех культурах фибробластов хромосома 1 относится, по данным всех исследователей, к группе поздних репликантов, а хромосома 2, как это точно установлено для первичной культуры эмбриональных фибробластов, заканчивает репликацию значительно ранее хромосомы 1 (С. И. Слезингер и А. А. Прокофьева-Бельговская, 1965, 1966). Хромосома 16 в культурах фибробластов ведет себя отчетливо как поздний редупликант. Данные о ее поведении в культуре лейкоцитов противоречивы, однако обстоятельные исследования относят хромосому 16 к ранним репликантам (Gilbert и др., 1962; С. И. Слезингер и А. А. Прокофьева-Бельговская, 1966, 1968). Хромосомы 4—5 в культуре лейкоцитов реплицируются позднее, чем в культуре эмбриональных фибробластов (рис. 52).
Очень вероятно, что обнаруженные различия в поведении хромосом при репродукции в культурах клеток разного типа, если они подтвердятся дальнейшими исследованиями, обусловлены различиями дифференциального функционирования этих хромосом в данных клетках (Slesinger, Ргоkofieva-Belgovskaya, 1968). Если по аналогии с Х-хромосомами женщины позднее окончание репликации ДНК и в аутосомах связано со спирализацией и генетической инактивацией участков хромосомы в периоде S клеточного цикла, то в отношении хромосом человека можно высказать следующее предположение: в культуре лейкоцитов генетически репрессированы участки хромосом 4—5 и относительно наиболее активны некоторые участки хромосом 1, 16 и 21—22, тогда как в культуре делящихся фибробластов интенсивно функционируют (кодирование m-РНК) участки хромосом 4—5 и генетически репрессированы некоторые участки хромосом 1, 16 и 21—22. Хромосомы 2, 3, 13—15, 17, 18 и 19—20 интенсивно функционируют во всех рассмотренных культурах.