Несмотря на большое разнообразие как форм опухолей, так и обнаруживаемых в их клетках изменений кариотипа, характер хромосомной изменчивости в разных опухолях имеет некоторые общие черты.
Прежде всего обращает на себя внимание тот факт, что в большинстве изученных опухолей кариотип клеток изменен по сравнению с нормальным диплоидным кариотипом, присущим данному виду животного или человека. Степень изменения бывает разной: от незначительного нарушения числа хромосом (гипердиплоидия или гиподиплоидия) до резкой полиплоидии (описаны случаи октаплоидных клеток и клеток с более высокой степенью плоидности в опухолях). Анэуплоидия в области диплоидных, триплоидных или тетраплоидных чисел является характерной. В недавних работах, содержащих суммарные данные разных авторов об изменении числа хромосом в опухолях человека (Yamada и др., 1966; Sandberg и др., 1967), показано, что истинно диплоидных опухолей у человека практически вообще не обнаружено (табл. 24 и 25). Опухоли, характеризующиеся 46 хромосомами, при более детальном анализе оказываются псевдодиплоидными.


У мышей и некоторых других животных описаны диплоидные опухоли. Однако не исключено, что это либо нераспознанные (в силу особенностей хромосомного набора) псевдо диплоидные опухоли, либо их кариотип содержит незначительные изменения, не выявляемые современными методами хромосомного анализа. Не исключено также, что некоторые опухоли животных имеют действительно нормальный кариотип, а генетическое изменение, связанное с малигнизацией, произошло в таких опухолях на молекулярногенном уровне.
Итак, первой чертой кариотипа большинства изученных опухолей является отклонение его от нормы. Второй характерной чертой является кариотипический полиморфизм опухолевых клеточных популяций. Клетки опухолей отличаются по кариотипу не только от нормальных соматических клеток, но и нередко друг от друга. Вариация числа хромосом в пределах одной и той же опухоли замечена давно, но лишь с разработкой современных методик изучения метафазных хромосом эта вариация смогла быть подвергнута более подробному анализу и количественной оценке. Оказалось, что в разных опухолях хромосомный полиморфизм выражен в различной степени. Есть опухоли, где большинство клеток обладает одинаковым числом хромосом, а есть и такие, особенно среди перевиваемых опухолей животных и человека, изучаемых обычно в далеко зашедших стадиях процесса,, где обнаруживается очень большое число клеточных вариантов. Большей частью в опухолях имеется преобладающий класс одинаковых по кариотипу клеток, составляющих на гистограмме модальный класс, и различное для разных опухолей число клеток — вариантов, отклоняющихся в обе стороны от модальных клеток. Клетки, составляющие в популяции большинство и сохраняющие особенности кариотипа в процессе роста и при пассировании опухолей, получили название стволовой линии (stemline), так как именно эти клетки обеспечивают рост опухоли и сохранение ее основных свойств (Hauschka, 1961; Makino, 1957).
На основании данных о числе хромосом клеток стволового класса, степени выраженности этого класса (процент таких клеток по отношению к общему числу изученных метафаз), пределов вариации числа хромосом и количества разных клеточных вариантов можно составить хромосомную характеристику каждой опухоли или клеточной популяции (Levan, 1959).
Кроме этих количественных показателей, хромосомная характеристика включает данные о наличии в кариотипе клеток стволового класса маркерных хромосом, возникающих вследствие структурных перестроек (транслокаций, инверсий, делеций) хромосом. На рис. 180—182 приведены примеры клеток некоторых новообразований, обладающих маркерными хромосомами. Наличие таких хромосом позволяет легко идентифицировать делящиеся опухолевые клетки среди нормальных клеток, что представляется важным методическим моментом как для диагностики раковых клеток (например, в асцитических и плевральных выпотах) (Ishiłiara и др., 1963), так и для изучения патогенеза злокачественных новообразований (Е. Е. Погосянц, Е. Л. Пригожина, А. А. Ставровская, 1966; А. А. Ставровская и Е. Л. Пригожина, 1967).
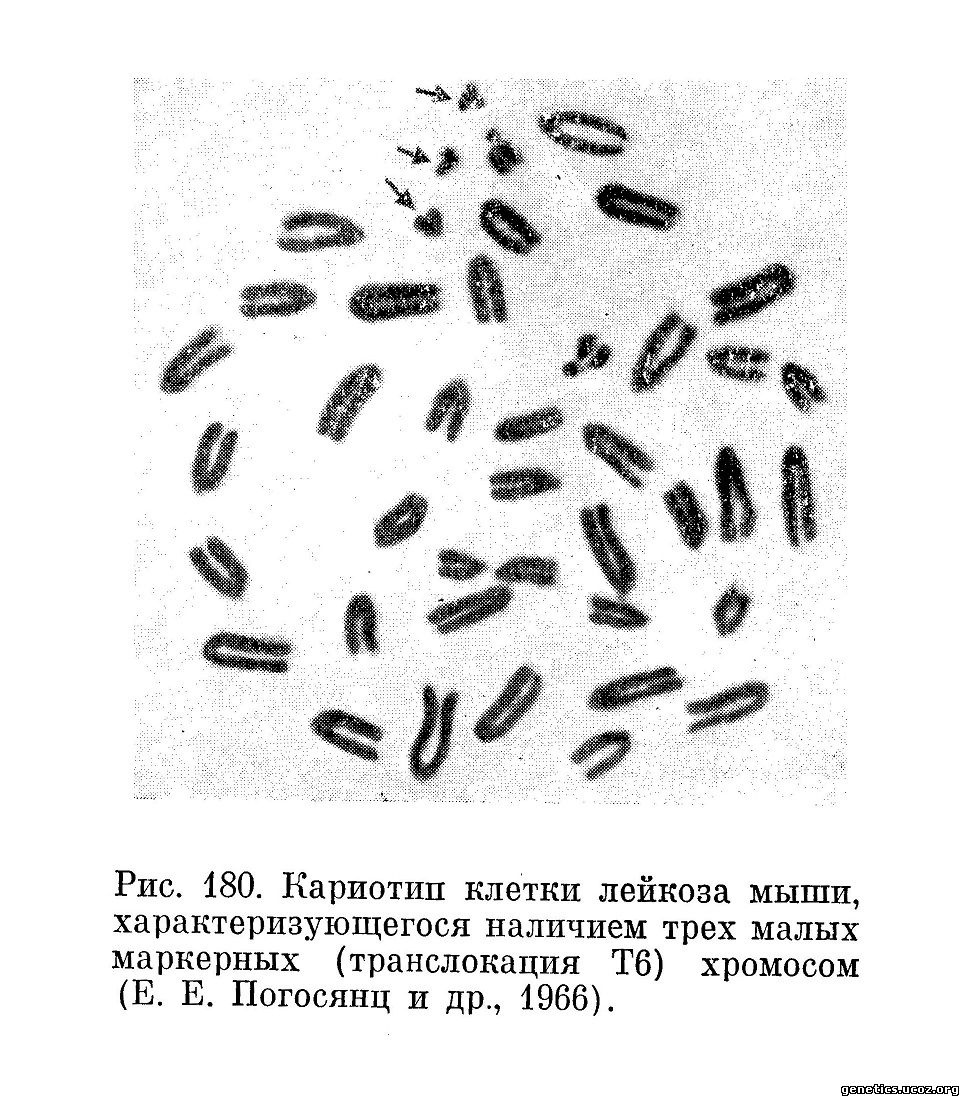
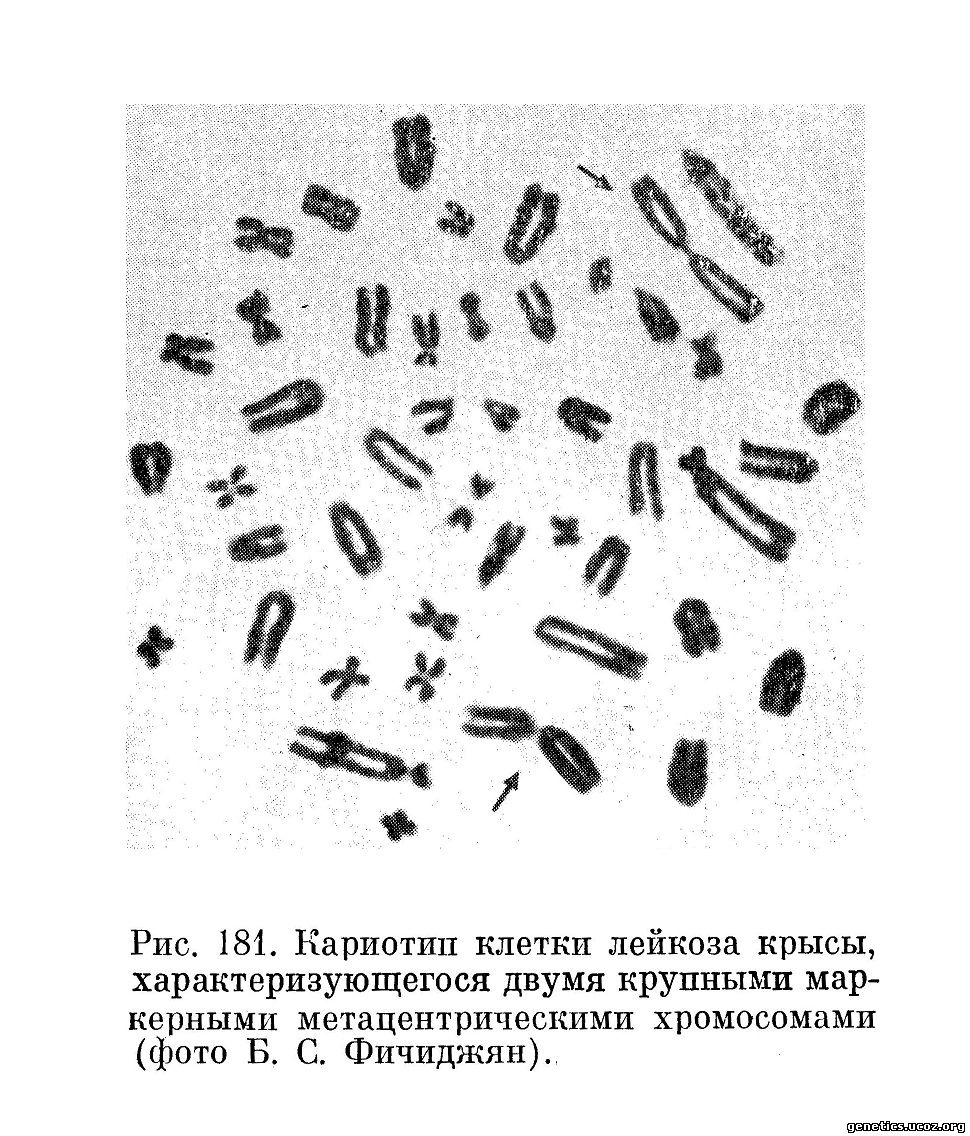
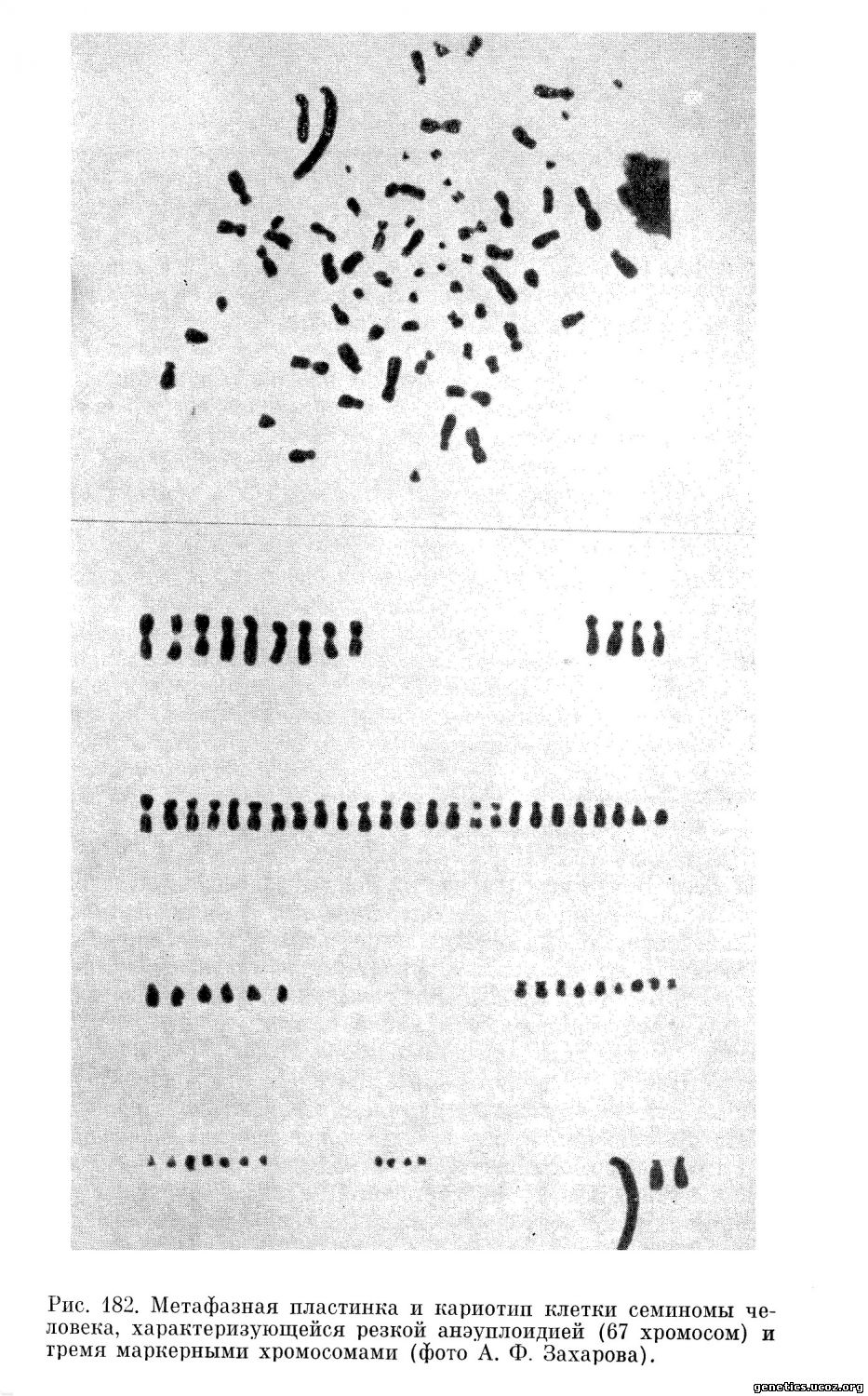
Иногда все или почти все клетки данной опухолевой популяции обладают одинаковым изменением кариотипа, содержат одну и ту же маркерную хромосому или даже набор маркерных хромосом. В таких случаях можно с уверенностью говорить о клоновом происхождении данной клеточной популяции (Б. С. Фичиджян и Е. Е. Погосянц, 1963; Atkin, Baker, 1966; Ford, Clark, 1963; Wakonig-Vaartaja, 1962), т. е. о том, что все ее члены являются потомками одной исходной измененной клетки. Это заключение исходит из невероятности предположения о повторном или параллельном возникновении идентичных сложных изменений кариотипа в разных клетках в процессе возникновения или роста новообразования. Все данные современной генетики о хромосомных аберрациях заставляют считать, что совладение одинакового изменения кариотипа во многих клетках данной популяции является результатом размножения одной исходной клетки, обладавшей таким изменением.
В некоторых случаях удается с большой долей вероятности предположить последовательность вторичных изменений кариотипа, возникавших за счет нерасхождения хромосом и их структурных перестроек в процессе размножения опухолевых клеток, т. е. составить картину динамики эволюции кариотипа данного новообразования на основе селекции определенных клеточных клонов (Lejeune и др., 1965).
Третьей чертой цитогенетической характеристики опухолей является ее относительная стабильность. Стабильность выражается в сохранении перечисленных выше количественных показателей и качественных особенностей кариотипа данной опухоли на протяжении многих ее пассажей у животных либо многих месяцев или лет роста в организме человека. Иногда при прогрессировании заболевания, даже после периода ремиссии, злокачественные клетки обладают тем же самым кариотипом, который был обнаружен до ремиссии (Ishihara и др., 1961; Sandberg и др., 1964; Reisman и др., 1964).
Наряду с такой кариотипической стабильностью описаны многочисленные случаи изменения хромосомной характеристики опухоли при изменениях условий ее пассирования и роста (В. К. Войтовицкий, 1965; Е. Е. По- госянц и А. Ф. Захаров, 1965; Б. С. Фичиджян, 1964; Hauschka, 1958). Эти перемены выражаются в сдвиге модального числа хромосом за счет как селективного размножения клеток, составлявших ранее в данной популяции небольшой процент, так и вновь возникающих кариотипических мутантов клеток. Процессы изменчивости и отбора в опухолевых клеточных популяциях удобно изучать на длительно перевиваемых клеточных культурах, являющихся моделью опухолевого роста (Hayflick, 1965). Изучение на протяжении большого числа пассажей характера и направления хромосомной изменчивости в клетках китайского хомячка — животного, обладающего сравнительно простым кариотипом, выявило интересную картину. Оказалось, что в анэуплоидных популяциях клетки с околодиплоидным кариотипом обладают селективными преимуществами по сравнению с крайними гипо-и гипердиплоидными вариантами. Иными словами, в таких популяциях отбор направлен в сторону достижения клетками максимальной кариотипической сбалансированности (Zakharov и др., 1964; А. Ф. Захаров и др., 1966; Е. С. Какпакова, 1966).
Каково биологическое значение кариотипической гетерогенности клеточных популяций опухолей и относительной стабильности хромосомных изменений?
Сохранение характера изменения кариотипа на протяжении многих сотен и тысяч клеточных поколений говорит о том, что оно не случайно и что клетки, обладающие таким кариотипом, отличаются явным селективным преимуществом не только перед нормальными клетками, но и перед другими вариантами опухолевых клеток в данной популяции. Интересно, что в то время как не только резкая анэуплоидия, но даже незначительные изменения числа и структуры хромосом, возникающие в половых клетках, сопровождаются весьма неблагоприятными для организма последствиями (вспомним хромосомные болезни), нарушения кариотипа в опухолевых клетках не только переносятся клеткой, но даже наделяют ее селективными преимуществами.
Это удивительное, можно сказать уникальное, явление стало понятным в результате многочисленных работ по цитогенетике злокачественных новообразований, проведенных за последнее десятилетие (см. обзоры Е. Е. Погосянц, 1963; Е. Е. Погосянц и А. Ф. Захаров, 1965; Hauschka, 1961; Levan, 1967).
Эти исследования показали, что хромосомная изменчивость имеет для опухоли приспособительное значение. Анэуплоидия и структурные нарушения кариотипа, приводя к сдвигам генного баланса клетки, как правило, сопровождаются изменением ее биологических свойств — химио-и радиочувствительности, иммунологических свойств и др. (Hauschka, 1958; Б. С. Фичиджян, 1964; В. К. Войтовицкий, 1965). И хотя специфической связи между характером изменения кариотипа и характером или степенью изменения биологических свойств клетки, по-видимому, не существует (О. Н. Петрова и др., 1967; Isaka, 1965; Yosida и др., 1967), значение хромосомной изменчивости состоит в том, что она наделяет опухолевые популяции более разнообразными генетическими потенциями по сравнению с таковыми нормальных тканей, характеризующихся, как известно, кариотипической однородностью. Вследствие хромосомной гетерогенности клеток опухолей они оказываются достаточно лабильными и выживают при неблагоприятном изменении окружающей среды, например при химио-и радиотерапии. Возникновение лекарственной (или лучевой) устойчивости опухолей связано с селективным выживанием и размножением клеток, резистентных к данному воздействию.
Клеточная изменчивость и наследственность, а также отбор и выживание наиболее приспособленных клеточных вариантов лежат в основе явления роста и развития — так называемой прогрессии (Foulds, 1958) злокачественных новообразований.