Первые наблюдения над строением и поведением хромосом в оогенезе у человека относятся к 30-м годам. Хотя эти исследования были направлены в основном на установление числа хромосом и системы половых хромосом (Evans, Swezy, 1931; Andres, Vogel, 1936), они пролили свет и на некоторые существенные временные отношения мейотических преобразований хромосом в онтогенезе.
Данные о строении и поведении хромосом человека в оогенезе менее обширны, чем данные о сперматогенезе. Наиболее существенные материалы получены в исследованиях Andres и Vogel (1936) и в современных работах Ohno с соавторами (1961, 1962). Эти материалы, главным образом результаты работы Ohno с сотрудниками, и использованы в приводимом изложении процесса оогенеза у человека.
Примордиальные половые клетки дифференцируются в оогонии в яичниках у 2-месячных эмбрионов. Эмбрионы 10—11 мм длиной уже имеют хорошо развитые недифференцированные гонады.
а) 3-месячные эмбрионы (рис. 73—1)
В гонадах женских эмбрионов этого возраста кора содержит рыхлую смесь оогоний и фолликулярных клеток (рис. 73), находящихся в периоде активного митотического цикла.

В более глубоких слоях некоторые половые клетки уже закончили дифференцировку из оогоний в ооциты и находятся в стадии лептотены и зиготены I мейотической профазы. Интерфазные ядра фолликулярных клеток, так же как полового эпителия, положительны в отношении тела полового хроматина, тогда как оогониальные клетки полового хроматина не содержат (рис. 74). В оогониальных метафазах присутствует 46 хромосом (рис. 75).


б) 4-месячные эмбрионы (рис. 73—2).
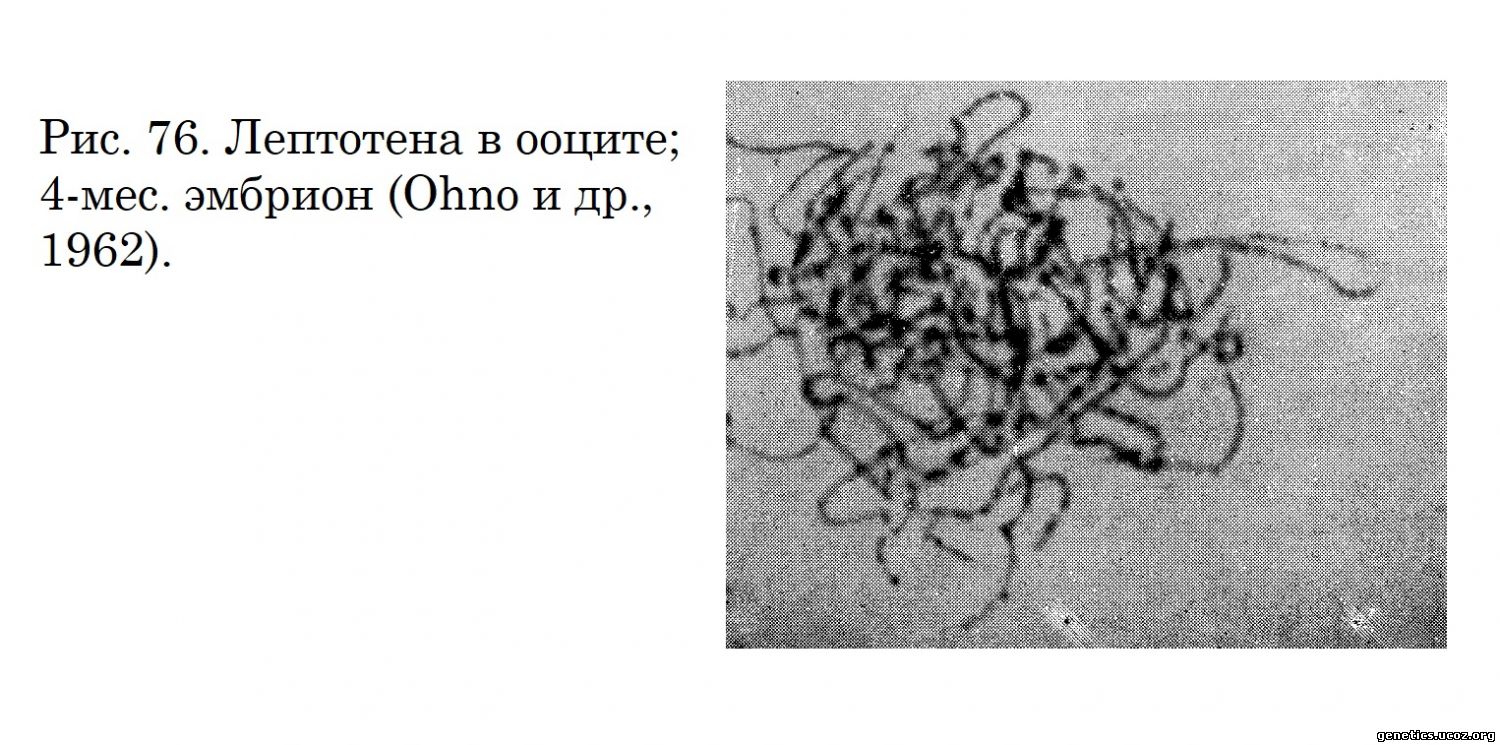
Область кортекса гонады сходна с предыдущим возрастом, она содержит оогонии и фолликулярные клетки, но в более глубоких слоях гонады наблюдается уже много ооцитов в стадии I мейотичеекой профазы. Многие клетки находятся в стадии лептотены и пахитены. Клетки, закончившие митотический цикл, имеют ядро, сходное по структуре с интерфазным, но больших размеров. Ядро в период лептотены содержит нежные нитевидные структуры, среди которых обе Х-хромосомы неразличимы вследствие их изопикнотичности с аутосомами (рис. 76).
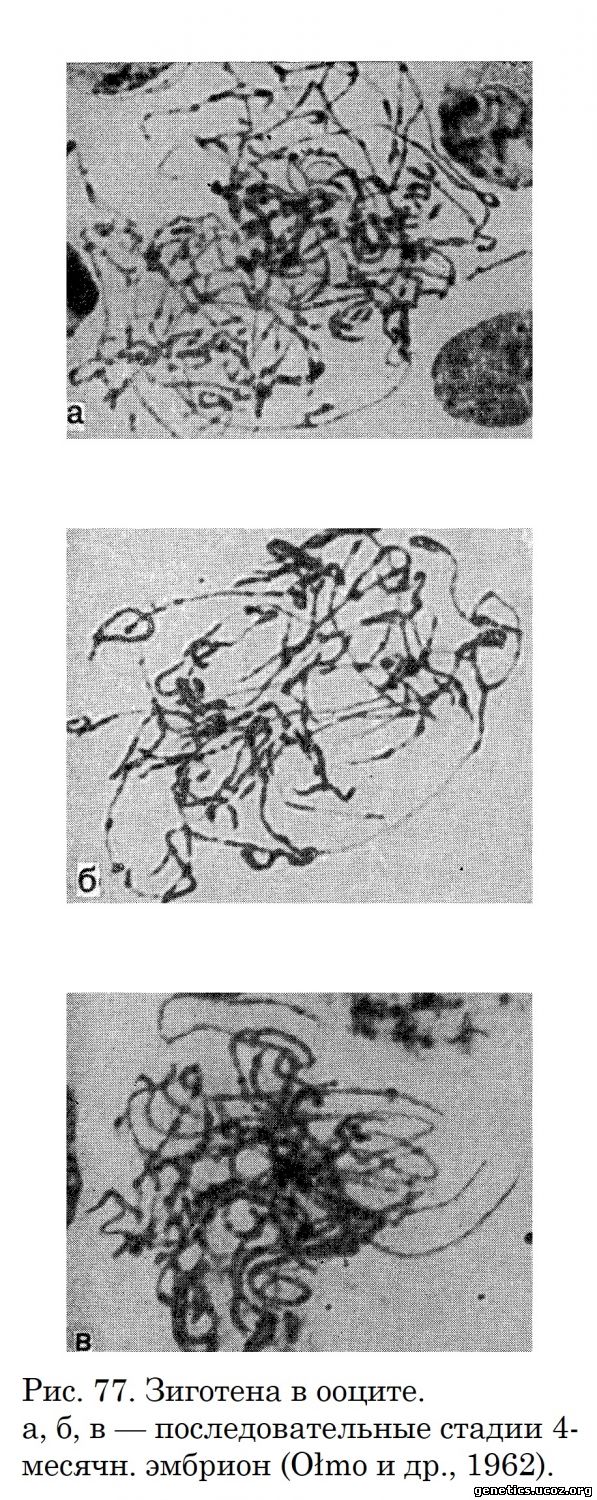
Зиготена в оогенезе занимает продолжительный период, в течение которого гомологичные хромосомы сдваиваются, постепенно спирализуются, укорачиваются и утолщаются (рис. 77). При этом в поздней зиготене часто формируется характерный для синапси с а плотный клубок хромосомных нитей (рис. 77, в). Половые хромосомы неотличимы от аутосом.
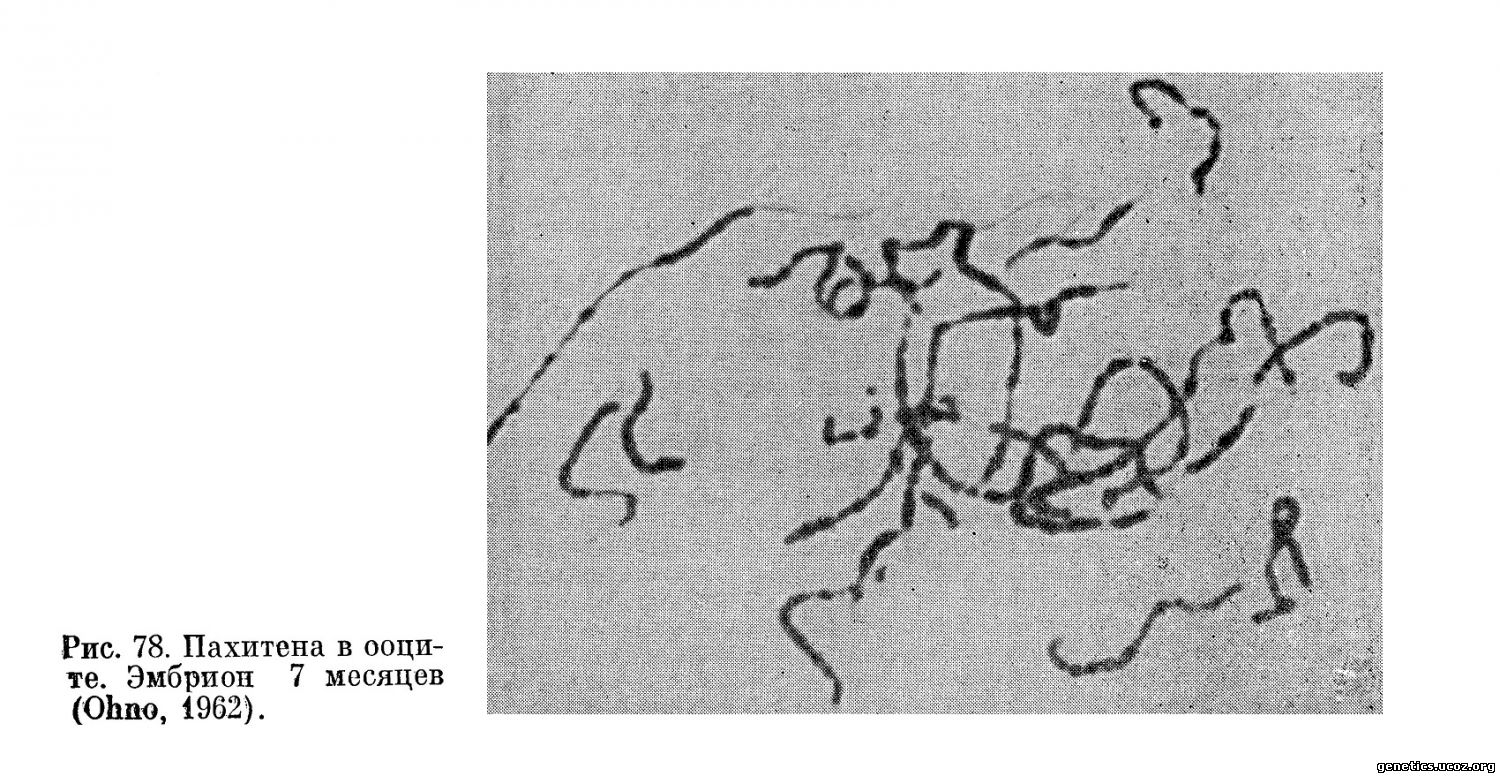
в) 7-месячные эмбрионы(рис. 73—3)
Поверхностный слой коры гонады не содержит оогоний; в нем обнаруживаются фолликулярные клетки в смеси с ооцитами, находящимися в стадии лептотены или зиготены. В более глубоких слоях коры каждый ооцит окружается компактным монослоем фолликулярных клеток; в его ядре продолжают развиваться пахитена и диплотена (рис. 78). Ядро входит в стадию пахитены, когда сдваивание гомологичных хромосом полностью закончено. Пахитенные нити имеют хорошо выраженную четковидную структуру.
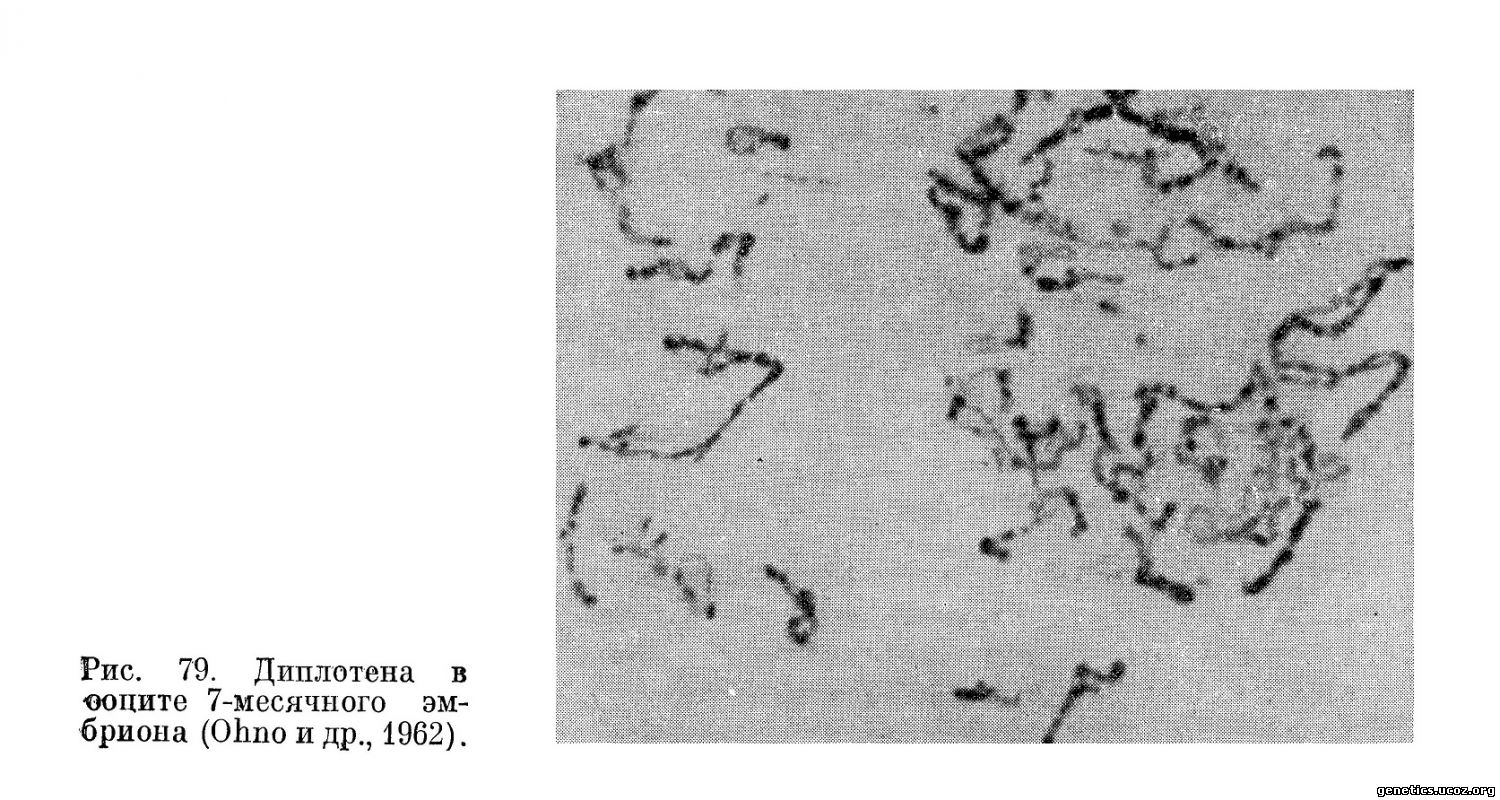
В диплотене происходит отталкивание гомологов, которые остаются соединенными хиазмами. В отличие от диплотенных бивалентов в сперматогенезе в оогенезе биваленты в дидлотене остаются в виде сильно удлиненных нитей, что затрудняет анализ хиазмообразования (рис. 79). В отличие от сперматогенеза диплотенные нити в оогенезе не претерпевают конденсации, но, наоборот, удлиняются и становятся неотличимыми друг от друга.
г) 9-месячные эмбрионы (рис. 73—4)
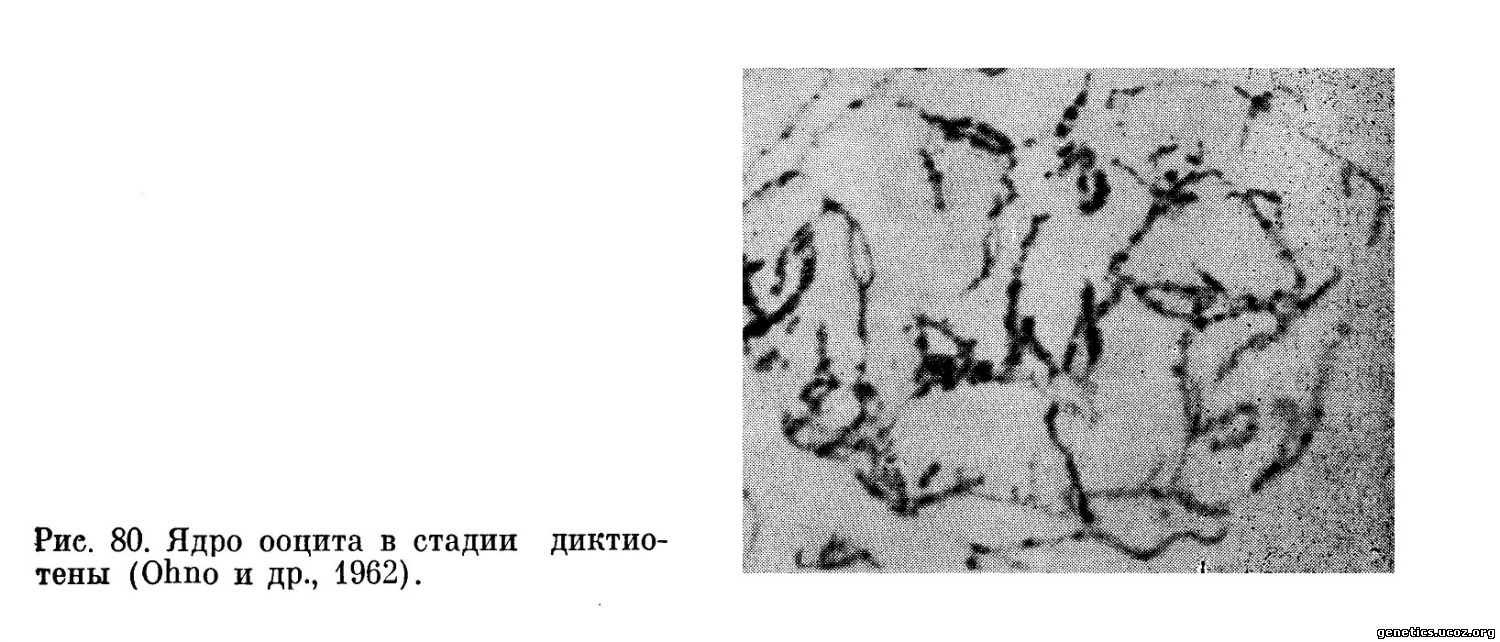
В яичнике эмбриона этого возраста находится до 400 000 фолликулов. Каждый фолликул содержит ооцит, окруженный плотным монослоем фолликулярных клеток. Ядро ооцита сходно с интерфазными структурами хотя в действительности оно содержит диплотенные биваленты, находящиеся в крайне вытянутом нитевидном состоянии. Эта стадия в оогенезе обозначается как диктиотена (рис. 80).
За несколько недель до рождения ядра всех ооцитов проходят через стадии лептотены, зиготены, пахитены и диплотены и входят в диктиотену. Они выходят из этой стадии только непосредственно перед первым мейотическим делением, когда фолликул вырастает до полных размеров, увеличиваясь в 80—90 раз, и раскрывается. Это происходит только по достижении женщиной половой зрелости. Второе мейотическое деление может наблюдаться в неоплодотворенном зрелом яйце, проходящем в фаллопиевы трубы.
В процессе роста фолликула и яйца увеличенное в размерах диктиотенное ядро яйцеклетки передвигается к периферии. Удлиненные нитевидные биваленты быстро конденсируются, хиазмы терминализируются и биваленты располагаются в метафазу первого мейотического деления. Благодаря периферическому положению диктиотенного ядра и направлению оси веретена это деление состоит в отделении от крупной яйцеклетки маленького цитоплазматического тельца с гаплоидным набором хромосом — первое полярное тело. После того как вторичный ооцит и первое полярное тело пройдут в фаллопиеву трубу, ядра обоих клеток быстро проходят вторую мейотическую профазу и приступают ко второму мейотическому делению.
Анафаза второго деления и выделение второго полярного тела, по-видимому, имеют место только после овуляции и оплодотворения. Так как первое полярное тело также претерпевает деление, оплодотворенное яйцо часто связано с тремя полярными телами. Неоплодотворенное яйцо отмирает.