Несмотря на многочисленные исследования действия вирусов на хромосомы человека и млекопитающих in vivo и in vitro, природа возникающих под действием этого агента разрывов хромосом до сих пор остается невыясненной. Предположительно можно указать на три фактора, являющихся ближайшей причиной разрывов хромосом:
1) непосредственное внедрение нуклеиновой кислоты вируса в хромосомы;
2) мутагенное действие вирусов;
3) мутагенное действие продуктов общего метаболизма нарушенного вирусом, в клетке.
Исследования первой возможности связаны со многими трудностями эксперимента, в котором используются вирусы с радиоактивной меткой и авторадиография. Возможность включения вируса в ДНК хромосом выяснялась в двух работах: с мечеными аденовирусами 12 и 2 в культуре лейкоцитов периферической крови человека (Nichols, 1966) и с меченым вирусом герпеса в диплоидных клетках китайского хомячка (Rapp, Hsu, 1963). Обе работы дали отрицательные результаты.
Анализу второго возможного механизма действия вирусов — как мутагенного агента — уделено наибольшее внимание. Работы в этой области направлены на сопоставление характера повреждений, вызванных, с одной стороны, вирусами и с другой — различными другими мутагенами, механизм действия которых достаточно изучен.
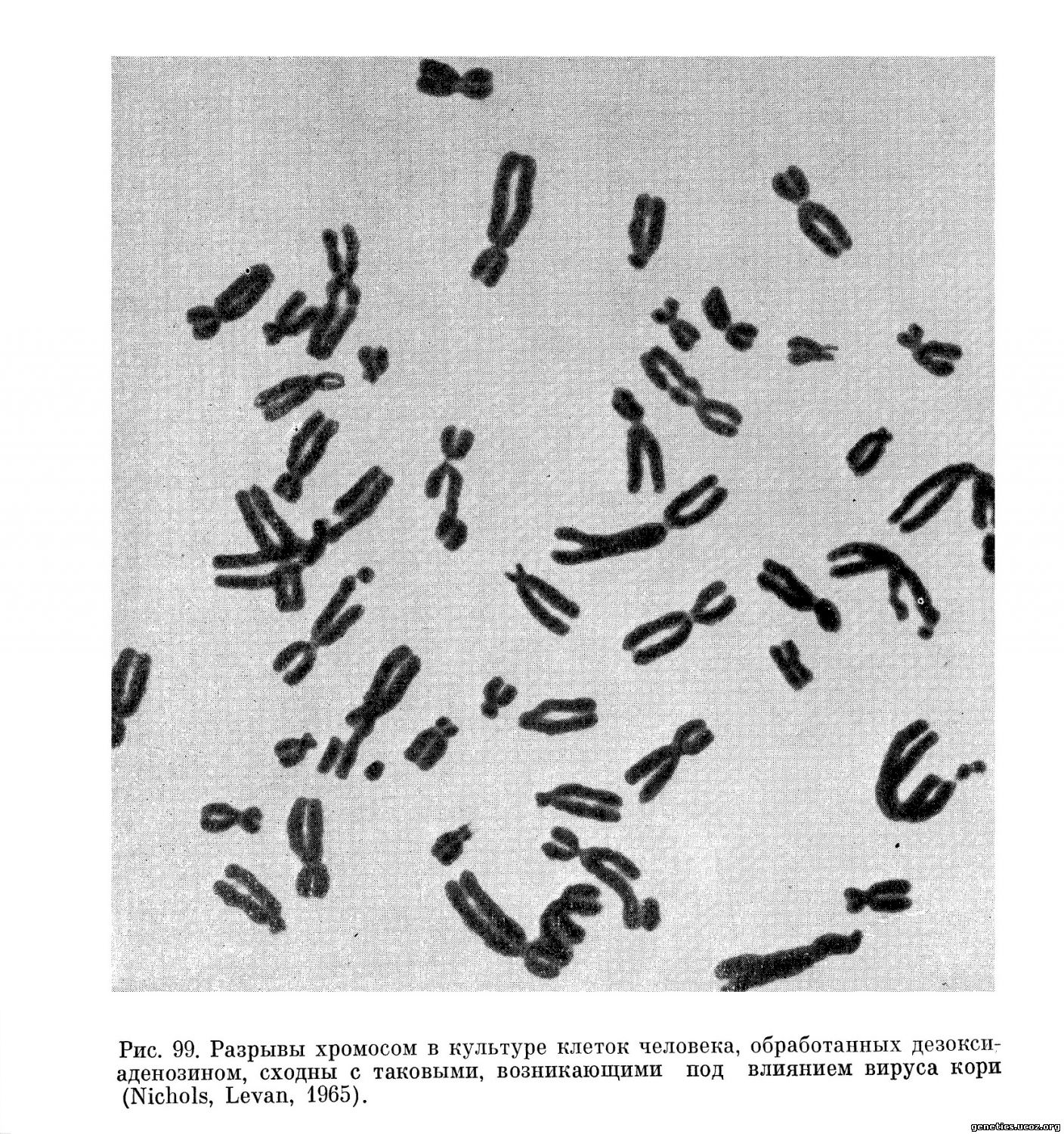
Обстоятельные исследования позволяют предполагать, что механизм мутагенного действия многих вирусов, стимулирующих разрывы хромосом, наиболее близок к механизму действия на хромосомы ряда ингибиторов синтеза ДНК: типы и локализация разрывов, вызванных вирусом герпеса, сходны с повреждениями хромосом, вызванными 5-бромдезоксиуридином {Stich, Hsu, 1963; Stich, Hsu, Rapp, 1964). Повреждения хромосом, индуцированные вирусами кори и ветряной оспы, подобны нарушениям, вызванным такими ингибиторами синтеза ДНК, как 5-фтордезоксиуридин или цитозин-арабинозид и дезоксиаденозин (рис. 98, 99) (Kihlman, 1963; Kihlman и др., 1963; Nichols и др., 1964).
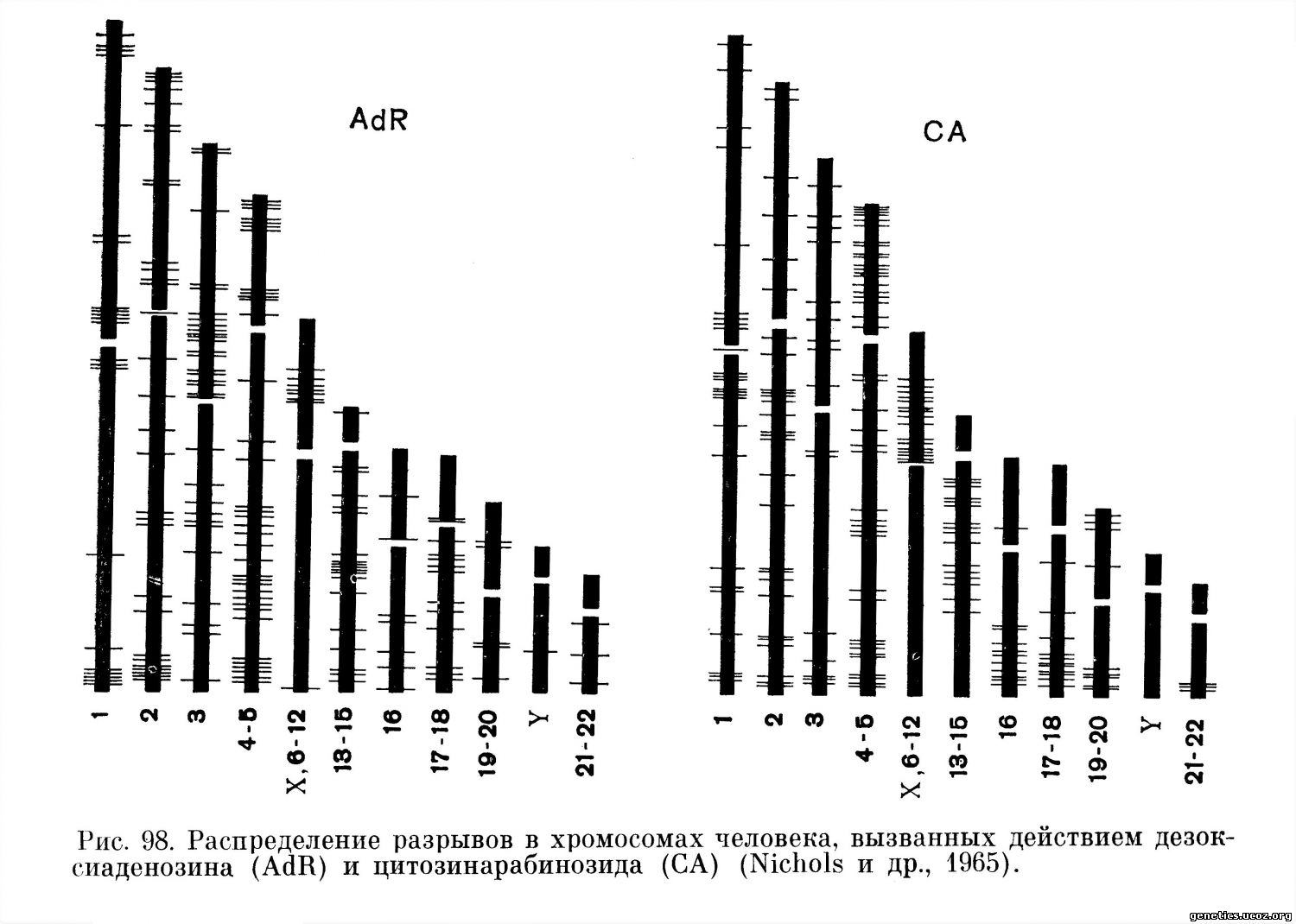
Распределение в хромосомах разрывов, индуцированных вирусом кори и обработанных дезоксиаденозином и цитозинарабинозидом, в высшей степени сходно: основная масса разрывов, индуцированных вирусом кори и этими ингибиторами синтеза ДНК, одинаково локализована в хромосомах X, 6—12 и 16.
Шведские исследователи показали, что вирус саркомы Рауса (штамм Шмидт—Руппина) и цитидинтрифосфат проявляют синергизм в индуцировании разрывов в хромосомах человека. При этом из 2000 локализованных разрывов в культуре лейкоцитов 17% разрывов, индуцированных этими мутагенами, проходят через центромерные районы хромосом (Nichols и др., 1965). Соответствие в локализации разрывов в хромосомах, индуцированных различными агентами, можно интерпретировать различно. Можно предполагать, что в хромосомах вообще содержатся некоторые «горячие» точки, особо чувствительные к различного рода воздействиям благодаря, например, наличию в них гетерохроматических участков (Kihlman, Levan, 1951; Moorhead, Saksela, 1963) или специфических пар оснований (Hsu, 1963). Можно сделать заключение, однако, и о сходстве самих механизмов действия этих агентов, к чему и склоняются шведские исследователи.
В пользу этого предположения приводятся следующие факты:
а) сходство морфологической картины повреждений хромосом и подавление обеими системами митотической активности;
б) способность и той, и другой системы очень быстро продуцировать дефекты хромосом после воздействия;
в) сходство в локализации разрывов по хромосомам;
г) синергизм действия вируса саркомы Рауса и цитидинтрифосфата в индуцировании разрывов хромосом у человека.
Механизм повреждающего действия на хромосому таких ингибиторов синтеза ДНК, как BUDR, согласно предположениям, связан с селективной заменой этим агентом тимина в молекуле ДНК. Если это так, то высокая частота разрывов в «горячих» точках хромосом во вторичных перетяжках, центромерных районах, гетерохроматических районах вообще может служить указанием на высокую концентрацию в этих районах пар оснований типа АТ.
Специфичность локализации разрывов в определенных хромосомах и определенных участках хромосом, возможно несколько различная при действии разных вирусов, может быть объяснена, таким образом, аналогичным же механизмом: на вирусную инфекцию первыми в ядре отвечают гетерохроматические районы хромосом (теломерные и центромерные районы, вторичные перетяжки). Можно высказать предположение о возможном значении в этой специфичности сходства состава ДНК гетерохроматических районов и вирусов (Hsu, Somers, 1961; Green, Pina, 1963).
Основным типом повреждений хромосом, возникающих при действии различных вирусов, являются хроматидные разрывы или хроматидные просветы.
Это дает основание заключить, что повреждение происходит в периоде S клеточного цикла при репродукции хромосомы. Таким образом, гипотеза о действии вирусов на хромосому как локальных ингибиторов синтеза ДНК достаточно хорошо обоснована.
Высказаны также предположения о роли токсических продуктов в повреждении хромосом в результате разрушения вирусом клетки или искажения ее метаболизма (Aula, 1965).
Gripenberg (1965) полагает, что широкая вариабельность повреждаемости хромосом при действии одного и того же вируса у разных пациентов указывает на то, что разрывы хромосом вызываются комплексированием целого ряда факторов.
Сопоставление локализации разрывов, вызванных действием вирусов, и спонтанных разрывов в контроле обнаружило, что оба типа разрывов проходят в одних и тех же участках. Поэтому сделано также заключение, что различия между опытом и контролем лишь количественные, а не качественные, т. е. вирусная инфекция лишь усиливает тенденцию, имеющую место в норме (Hampar, Ellison, 1961, 1963).
Evans (1967) выдвигает весьма стимулирующую гипотезу для объяснения природы численных нарушений хромосомных комплексов в результате действия вирусов. Он полагает, что одним из важнейших результатов действия вируса на клетку являются изменения в поведении ядрышка — оно во многих случаях резко увеличивается в размерах и длительно сохраняется в митозе.
Именно это обстоятельство, по мнению Evans, может иметь решающее значение в нарушениях нормального поведения хромосом в оогенезе, особенно если учесть длительность диктиотенной стадии, и сопровождаться формированием анэуплоидных гамет. Задержанная резорбция ядрышка, пораженного вирусом, может резко затруднить нормальное распределение хроматид между полюсами в митозе и гомологичных хромосом в мейозе. В пользу этого представления свидетельствуют факты наиболее частой анэуплоидии при вирусных инфекциях по ядрышкообразующим хромосомам.